An
activation of parvocellular oxytocinergic
neurons in the paraventricular nucleus in
oxytocin-induced yawning and penile
erection
Kita I, Yoshida Y, Nishino S.
Sleep and Circadian
Neurobiology Laboratory, Center for Narcolepsy,
Department of Psychiatry and Behavioral
Sciences, Stanford University. USA
-Kita I, Kubota N,
Yanagita S, Motoki C Intracerebroventricular
administration of corticotropin-releasing factor
antagonist attenuates arousal response
accompanied by yawning behavior in rats.
Neurosci.Letter 2008;433(3):205-208
-Kita I, Yoshida
Y, Nishino S. An activation of parvocellular
oxytocinergic neurons in the paraventricular
nucleus in oxytocin-induced yawning and penile
erection. Neurosci Res. 2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Behavioural Brain Research
2000;117(1-2):119-126
-Kubota N, Amemiya
S, Motoki C, Otsuka T, Nishijima T, Kita I.
Corticotropin-releasing factor antagonist
reduces activation of noradrenalin and serotonin
neurons in the locus coeruleus and dorsal raphe
in the arousal response accompanied by yawning
behavior in rats. Neurosci Res.
2012;72(4):316-323
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Sato-Suzuki I,
Kita I, Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5)2765-2775
Intracerebroventricular (ICV) or PVN local
injections of oxytocin induce yawning and penile
erection, for which a positive feedback
mechanism for the PVN oxytocinergic activation
is suggested, but this had not been directly
substantiated in vivo.
We have and significantly increased the
percentage of c-Fos positive oxytocin neurons in
the medial, dorsal and lateral parvocellular
subdivision of the PVN. However, increases in
the magnocellular portion were not significant.
We also found that lithium chloride (LiCl, 0.5
and l.0mEq), a compound known to activate
oxytocinergic neurons, also significantly
increased the percentage of c-Fos positive
oxytocin neurons in all PVN portions. However,
LiCl did not induce yawning and penile erection,
but counteracted the oxytocin-induced yawning
and penile erection. These results suggest that
if the activation of oxytocinergic neurons in
the PVN is important for mediating
oxytocin-induced yawning and penile erection, a
selective activation of parvocellular
oxytocinergic neurons in the PVN is likely to be
involved.
1. Introduction
A series of animal studies has demonstrated
that the oxytocinergic paraventricular nucleus
(PVN) in the hypothalamus is involved in sexual
behavior (Witt and Insel, 1994; Meus et al.,
1999; Argiolas and Meus, 2004; Nishitani et al.,
2004). In male rats, the number of activated
oxytocinergic neurons that express c-Fos
(protein product of an immediate early gene,
c-fos) in the PVN was higher in sexually
experienced animals (Witt and Insel, 1994;
Nishitani et al., 2004).
Intracerebroventricular (1CV) and/or local
administration of oxytocin, dopamine D2 receptor
agonists, adrenocorticotropin (ACTH) and
alpha-melanocyte stimulating hormone (a-MS H)
into the PVN induced penile erection and yawning
in male rats (Argiolas et al., 1985; Argiolas et
al., 1988; Fujikawa et al., 1995; Argiolas et
ai., 2000), and it is also believed that an
activation of the descending oxytocinergic
system in the PVN is one of the most important
mechanisms for mediating the induction of penile
erection and yawning by these pharmacological
manipulations (Argiolas et ai, 1987b; Meus et
al., 1989; Argiolas et ai, 1993; Pandita et al.,
1998; Tang et al., 1998; Veronneau-Longueville
et al., 1999; Andersson, 2001).
The oxytocinergic system in the PVN can be
divided into magnocellular and parvocellular
neurons (Swanson and Kuypers, 1980; Sawchenko
and Swanson, 1982; Swanon and Sawchenko, 1983).
It has been reported that parvocellular
oxytocinergic neurons (especially in the caudal
part of the parvocellular division of the PVN)
send descending axons to the lower brain stem,
including the locus coeruleus, nucleus of the
solitary tract, the dorsal motor nucleus of the
vagus nerve, ventrolateral parts of the
reticular formation and the spinal cord (Swanson
and McKellar, 1979; Swanson and Sawchenko, 1983;
Wagner and Clemens, 1991). On the other hand,
magnocellular oxytocinergic neurons in the PVN
mainly synthesize the neurohypophyseal hormone
oxytocin and send their axons into the posterior
pituitary gland (Brownstein et al., 1980; Rhodes
et al., 1981; Swanson and Sawchenko, 1983).
Witt and Insel (1994) previously had
demonstrated in a c-Fos study that a
preferential activation of the parvocellular
regions of the PVN is involved in the copulatory
interactions of male rats. Similarly, several
authors suggested that the descending
parvocellular oxytocinergic neurons in the PVN
could be specifically involved in penile
erection and yawning related responses such as
arousal, respiratory, and cardiovascular
responses after the above-mentioned
pharmacological manipulations (Wagner and
Clemens, 1991; Argiolas and Melis, 1998;
Sato-Suzuki et al., 1998; Andersson, 2001).
Particularly, an involvement of a positive
feedback loop of the PVN oxytocinergic system
for oxytocin-induced penile erection was
proposed (Melis et al., 1986; Argiolas and
Melis, 1998; Andersson, 2001).
In the present study, we have investigated
the involvement of parvocellular oxytocinergic
neurons in the induction of penile erection and
yawning in vivo with double-staining for c-Fos
and oxytocin in the PVN after ICV oxytocin
administration. We found that preferential
activation of parvocellular oxytocinergic
neurons in the PVN is associated with
oxytocin-induced penile erection and
yawning.
4. Discussion
Functional anatomical assessments of PVN
oxytocinergic neurons in the current study have
suggested that a selective activation of
parvocellular oxytocinergic neurons in the PVN
of the hypothalamus is associated with
oxytocin-induced penile erection and
yawning.
Although centrally-administered oxytocin may
diffuse widely and activate various oxytocin
receptive sites to induce penile erection and
yawning, a series of animal studies (Melis et
al., 1986; Argiolas et al., 1987a; Melis et al.,
1987; SatoSuzuki et al., 1998; Kita et al.,
2000; Zahran et al., 2000) has suggested that
the PVN of the hypothalamus is one of the most
important brain structures in oxytocin-induced
penile erection and yawning. Melis et al. (1986)
showed that a micro-injection of oxytocin into
the PVN or into the CAI field of the hippocampus
induces a dose-dependent increase in the number
of penile erection and yawning episodes in male
rats, and that such a response was not observed
when oxytocin was injected into other brain
areas. It has been also shown that electrolytic
lesions of the PVN prevent yawning and penile
erection induced by 1CV injection of oxytocin
(Argiolas et al., l987a; Liu et al., 1997).
Involvement of the PVN oxytocin neurons for
copulatory interactions was also demonstrated by
several authors (Melis et al., 1999; Argiolas
and Melis, 2004; Nishitani et al., 2004), and an
induction of the c-Fos was observed in PVN
oxytocin neurons in sexually-experienced male
rats in the study by Witt and Insel (1994).
Exogenously administered oxytocin is likely
to stimulate oxytocin receptors to induce penile
erection and yawning. Oxytocin binding sites
have been demonstrated in the rat PVN (Brinton
et al., 1984; Freund-Mercier et al., 1994), and
1CV or local injection into the PVN of a potent
oxytocin antagonist,
d(CH2)5-Tyr(Me)-Orn8-vasotocin, were shown to
prevent penile erection and yawning induced by
oxytocin (Argiolas et al., 1987b; Meus et al.,
1989; Meus et al., 1992; Argiolas et al., 1993;
Melis et al., 1999). These results suggest that
oxytocin-induced penile erection and yawning are
mediated by the PVN oxytocin receptive
sites.
A positive feedback control of the oxytocin
system in certain physiological functions, such
as milk ejection, is known: oxytocinergic
neurons are activated by physiological stimuli
such as suckling (Freund-Mercier and Richard,
1984). Since several experiments have shown that
oxytocin applications enhance the electrical
activity of oxytocinergic neurons
(Freund-Mercier and Richard, 1981; Jourdain et
al., 1998) and stimulate the release of oxytocin
(Moos et al., 1984), and since
oxytocin-immunoreactive synapses have been found
to impinge on oxytocinergic neurons in the
hypothalamic nuclei (Theodosis, 1985), several
authors (Meus et al., 1986; Argiolas and Meus,
1998; Andersson, 2001) have also argued the
possibility that oxytocin activates
oxytocinergic neuron in the PVN to induce penile
erection and yawning through a similar pathway
that is involved during copulatory behaviors
(Argiolas and Melis, 2004).
In the current study, we demonstrated that
an ICV injection of oxytocin significantly
increases c-Fos positive oxytocinergic neurons
in the mp, dp, and lp subdivisions of the PVN,
and induced yawning and penile erection. The
magnitude of increase in c-Fos expressions in
these structures after ICV oxytocin injection
was small, and not more than 10% of oxytocin
positive neurons in these structured are
activated.
These results suggest that non-oxytocin
neurons such as corticotoropin releasing factor
(CRF) neurons are also activated during yawning
and penile erection. However, it is not known
whether all oxytocin neurons are stained with
the antibody we used, or whether a large number
of non-oxytocin ergic neurons are indeed
significantly activated. Similarly, it is not
known whether these short-lasting phasic events
(i.e. penile erection and yawning) are mediated
by tonic and strong activation of oxytocinergic
neurons. Interestingly, however, oxytocinergic
neuron activation with oxytocin injection is
rather selective for the parvocellular regions,
and no significant increase in c-Fos expression
in the magnocellular oxytocinergic neurons were
observed.
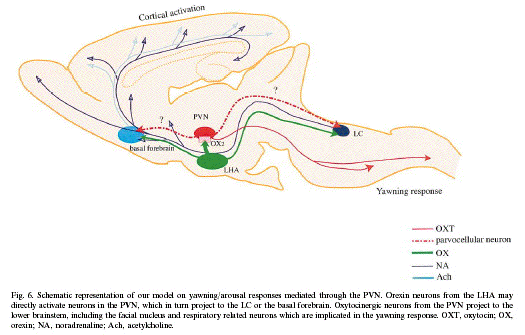
Parvocellular oxytocinergic cells in the
caudal portion of the PVN send descending axons
to the lower brain stem, including the locus
coeruleus, nucleus of the solitary tract, the
dorsal motor nucleus of the vagus nerve,
ventrolateral medulla and the spinal cord
(Swanson and McKellar, 1979; Sawchenko and
Swanson, 1982; Wagner and Clemens, 1991; Tang et
al., 1998). The projections to the spinal cord
include those to the spinal autonomic structures
innervating the penis, and to the ventral horn
motoneurons in the lumbar spinal cord that
innervates the bulbospongiosus of the penis
(Schmidt, 2005). Oxytocin binding sites are also
reported in these projection sites (Tribollet et
al., 1997). The PVN lesion causes an almost
complete depletion of oxytocin in brainstem and
spinal cord (Lang et al., 1983; Argiolas et al.,
1987a; Ackerman et al., 1997; Liu et al., 1997).
These oxytocinergic efferent brainstem regions
are involved in arousal, respiratory,
cardiovascular, and other autonomic functions
including penile erection and yawning. Our
results are consistent with the results of these
anatomical findings and both findings indicate
the importance of parvocellular oxytocinergic
neurons in the PVN for the induction of penile
erection and yawning by ICV oxytocin injection.
Preferential activation of oxytocin neurons in
the parvocellular regions (over the
magnocellular regions) were also observed in the
induction of c-Fos in sexually-experienced male
rats, and the results further suggest that
oxytocin administration may activate the pathway
involved in physiological copulatory
behaviors.
Several studies (Verbalis et al., 1986;
O'Connor et al., 1987; McCann et al., 1989; Cui
et al., 2001; Wirth et al., 2002) have suggested
that intraperitoneal injection of LiCl increases
the activity of oxytocin neurons and rises in
the plasma concentration of oxytocin. In
agreement with previous studies (Portillo et
al., 1998; Cui et al., 2001), we also have
observed that systemic administration of LiCl
induces c-Fos expression in both PVN
magnocellular oxytocinergic and parvocellular
oxytocinergic neurons.
However, LiCl (0.5 and 1.0 mEq, i.p.) did
not induce any yawning or penile erection, but
rather counteracted the oxytocininduced yawning
and penile erection. These results are difficult
to reconcile with the hypothesis that activation
of the PVN oxytocinergic neurons are involved in
oxytocin-induced yawning and penile erection.
Our c-Fos results have shown that the magnitude
of the activation of oxytocinergic neurons by
LiCl is much higher than those by ICV oxytocin.
Moreover, LiC1 administration significantly
increased c-Fos positive oxytocin neurons in all
PVN portions, including magnocellular
oxytocinergic neurons. This may also be
consistent with the finding that intraperitoneal
injection of LiCl significantly increases plasma
toncentration of oxytocin, since magnocellular
oxytocinergic neurons send their axons into the
posterior pituitary gland to secrete oxytocin
into the systemic circulation.
It is therefore possible that a strong
activation of oxytocin neurons in the whole
portion of PVN by systemic LiCl administration
might mask the effects of activation of
parvocellular oxytocinergic neurons that are
more specifically related to the induction of
penile erection and yawning. Since it is also
likely that both oxytocin and LiCl also activate
non-oxytocin neurons in the PVN, activation of
these neurons may also play a role in
oxytocin-induced penile erection and yawning. In
addition, dendritic secretion of oxytocin
(especially from the magnocellular oxytocinergic
neurons) has also been reported and this also
involves the sexual behavior (Sabatier et al.,
2003). Therefore, more complex interactions may
be considered for explaining the results
observed in the current study.
In summary, we observed that ICV oxytocin
injection in male rats induced yawning and
penile erection and selectively increased the
expression of c-Fos in the parvocellular
oxytocinergic neurons. On the other hand,
although systemic administration of LiCl that
potently activated PVN oxytocin neurons in the
all divisions of the PVN, it did not induce
yawning and penile erections, but rather
counteracted the oxytocin-induced yawning and
penile erection. These results suggest that if
an activation of oxytocinergic PVN neurons is
critical for the oxytocin-induced yawning and
penile erection, a selective activation of the
parvocellular oxytocinergic neurons are likely
to be involved. Our results also propose that
the double-staining of c-Fos and oxytocin
neurons in the PVN are useful in determining the
functional anatomical pathways for the penile
erection and yawning induced by various classes
of drugs (i.e. D2 receptor agonists, ACTH and
a-MSH).
-Collins GT, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior. J Pharmacol Exp Ther
2005;314(1):310-319.
-Collins GT, Newman
AH,Woods JH et al.Yawning and hypothermia in
rats: effects of dopamine D3 and D2 agonists and
antagonists. Psychopharmacology
(Berl).
2007;193(2):159-170
-Collins GT. et
al. Food restriction alters
pramipexole-induced yawning, hypothermia, and
locomotor activity in rats: Evidence for
sensitization of dopamine D2 receptor-mediated
effects. JEPT 2008;325:691-697
-Collins
GT et al. Narrowing in on compulsions:
dopamine receptor functions Exp Clin
Psychopharmacol 2008,16(4):498-502
-Collins
GT et al. Pro-erectile Effects of Dopamine
D2-like Agonists are Mediated by the D3 Receptor
in Rats and Mice JPEP 2009;329(1):210-217
-Collins GT,
Truong YN, et al. Behavioral sensitization
to cocaine in rats: evidence for temporal
differences in dopamine D(3) and D (2) receptor
sensitivity. Psychopharmacology (Berl).
2011;215(4):609-620.
-Hipolide DC;
Lobo LL; De Medeiros R; Neumann B; Tufik S
Treatment with dexamethasone alters yawning
behavior induced by cholinergic but not
dopaminergic agonist. Physiol Behav
1999;65(4-5):829-832
-Hipolide
DC, Tufik S Paradoxical sleep deprivation in
female rats alters drug-induced behaviors
Physiol Behav. 1995;57(6):1139-1143
-Kita I, Kubota N,
Yanagita S, Motoki C Intracerebroventricular
administration of corticotropin-releasing factor
antagonist attenuates arousal response
accompanied by yawning behavior in rats.
Neurosci.Letter 2008;433(3):205-208
-Kita I, Yoshida
Y, Nishino S. An activation of parvocellular
oxytocinergic neurons in the paraventricular
nucleus in oxytocin-induced yawning and penile
erection. Neurosci Res. 2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Beh Brain Res
2000;117(1-2):119-126
-Kubota N, Amemiya
S, Motoki C, Otsuka T, Nishijima T, Kita I.
Corticotropin-releasing factor antagonist
reduces activation of noradrenalin and serotonin
neurons in the locus coeruleus and dorsal raphe
in the arousal response accompanied by yawning
behavior in rats. Neurosci Res. 2012
-Moyaho
A, Valencia J Grooming and yawning trace
adjustment to unfamiliar environments in
laboratory Sprague-Dawley rats J Comp Psychol
2002;116(3):263-269
-Neumann BG,
Troncone LR, Braz S, Tufik S Modifications
on dopaminergic and cholinergic systems induced
by the water tank technique: analysis through
yawning behavior. Arch Int Pharmacodyn Ther
1990;308:32-38
-Sato-Suzuki I,
Kita I; Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5):2765-2775
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Tufik S et
al Effects of stress on drug induced yawning
Physiol Behav 1995;58(1):1881-1884