Corticotropin-releasing
factor antagonist reduces activation of
noradrenalin and serotonin neurons in the locus
coeruleus and dorsal raphe in the arousal
response accompanied by yawning behavior in
rats
Kubota N, Amemiya S, Motoki C, Otsuka T,
Nishijima T, Kita I.
Department of
Human Health Science, Tokyo Metropolitan
University Japan.
-Kita I,
Kubota N, Yanagita S, Motoki C
Intracerebroventricular administration of
corticotropin-releasing factor antagonist
attenuates arousal response accompanied by
yawning behavior in rats. Neurosci.Letter
2008;433(3):205-208
-Kita I, Yoshida
Y, Nishino S. An activation of parvocellular
oxytocinergic neurons in the paraventricular
nucleus in oxytocin-induced yawning and penile
erection. Neurosci Res. 2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Behavioural Brain Research
2000;117(1-2):119-126
-Kubota N, Amemiya
S, Motoki C, Otsuka T, Nishijima T, Kita I.
Corticotropin-releasing factor antagonist
reduces activation of noradrenalin and serotonin
neurons in the locus coeruleus and dorsal raphe
in the arousal response accompanied by yawning
behavior in rats. Neurosci Res.
2012;72(4):316-323
-Kubota
N, Amemiya S, Yanagita S, Kita I. Neural
pathways from the central nucleus of the
amygdala to the paraventricular nucleus of the
hypothalamus are involved in induction of
yawning behavior due to emotional stress in
rats. Behav Brain Res. 2022 Sep 1:114091.
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Sato-Suzuki I,
Kita I, Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5)2765-2775
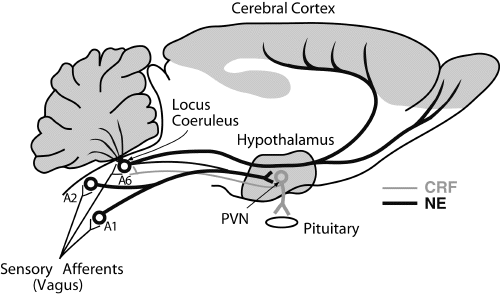
We previously reported that
intracerebroventricular (icv) administration of
corticotropin-releasing factor (CRF) antagonist
attenuates the arousal response during yawning
behavior in rats. However, the CRF-related
pathway involved in the arousal response during
yawning is still unclear. In the present study,
we assessed the involvement of the
CRF-containing pathway from the hypothalamic
paraventricular nucleus (PVN) to the locus
coeruleus (LC) and the dorsal raphe nucleus
(DRN) in the arousal response during frequent
spontaneous yawning, which was induced by
several microinjections of l-glutamate into the
PVN in anesthetized rats, using c-Fos
immunohistochemistry. The PVN stimulation showed
significant increases in activation of PVN CRF
neurons, LC noradrenalin (NA) neurons and DRN
serotonin (5-HT) neurons as well as arousal
response during yawning. But icv administration
of a CRF receptor antagonist, ?-helical CRF
(9-41), significantly inhibited the activation
of both LC NA neurons and DRN 5-HT neurons
except the activation of CRF neurons in the PVN,
and significantly suppressed the arousal
response during yawning. These results suggest
that the CRF-containing pathway from PVN CRF
neurons to LC NA neurons and DRN 5-HT neurons
can be involved in the arousal response during
yawning behavior.
1. Introduction
Yawning behavior is typically associated
with the induction of an arousal response (Concu
et al., 1974; Sato-Suzuki et al., 1998). We have
reported that the arousal response during
yawning can be evoked by several forms of
chemical stimulation of paraventricular nucleus
(PVN) in the hypothalamus in anesthetized,
spontaneously breathing rats, which was
characterized an arousal shift in the
electrocorticogram (EC0G), represented by lower
voltage and faster rhythms occurring prior to
yawning behavior (Kita et al., 2008; Sato-Suzuki
et al., 1998, 2002). Previous studies have
suggested that activation of oxytocin neurons
and corticotropin-releasing factor (CRF) neurons
in the PVN mediate the induction of the yawning
accompanied by arousal response (Argiolas et
al., 1987; Argiolas and Melis, 1998; Kita et
al., 2006a,b), and the CRF neurons in the PVN
could be primarily responsible for the arousal
response during yawning behavior (Dunn and
Berridge, 1990; Heinrichs and Koob, 2004; Kita
et al., 2008; Menzaghi et al., 1993). We
previously showed that intracerebroventricular
(icy) administration of a CRF antagonist
significantly attenuates the arousal response
during yawning induced by PVN stimulation (Kita
et al., 2008). However, the CRF-related pathway
responsible for the arousal response during
yawning behavior is still unknown. A series of
animal studies has suggested that CRF has
excitatory effects on the locus coeruleus (LC)
noradrenalin (NA) neurons and the dorsal raphe
nucleus (DRN) serotonin (5-HT) neurons, which
are the regions within the brainstem reticular
formation responsible for arousal response
(Curtis et al., 2002; jedema and Grace, 2004;
Kirby et al., 2000; Lowry et al., 2000; Schulz
and Lehnert, 1996; Smagin et al., 1995; Staub et
al., 2005, 2006). Taken together, the neural
projection from PVN CRF neurons to LC NA neurons
and DRN 5-HT neurons may be one of candidates
for the CRF-containing pathway responsible for
the arousal response during yawning. In the
present study, to investigate the involvement of
the CRFcontaining pathway in the arousal
response during yawning, we evaluated the effect
of CRF antagonist, ct-helical CRF (9-41) on
activation of LC NA neurons, DRN 5-HT neurons
and PVN CRF neurons as well as the arousal
response during yawning in anesthetized,
spontaneously breathing rats, using c-Fos
immunohistochemistry. Our
findings suggest that the CRF-containing
pathway from PVN CRF neurons to LC NA neurons
and DRN 5-HT neurons can be involved in the
arousal response during yawning behavior.
4. Discussion
Administration of CRF receptor antagonist
(ct-helical CRF (941)) into the ventricle
significantly suppressed the arousal response
during yawning, and significantly inhibited both
increases in activity of LC NA neurons and DRN
5-HT neurons during yawning induced by PVN
stimulation, but not activity of PVN CRF
neurons. These results suggest that CRF neurons
in the PVN may mediate the arousal response
during yawning behavior through activation of LC
NA neurons and DRN 5-HT neurons. This is the
first study suggesting the involvement of the
CRF-containing pathway from PVN CRF neurons to
LC NA neurons and DRN 5-HT neurons in the
arousal response during yawning behavior.
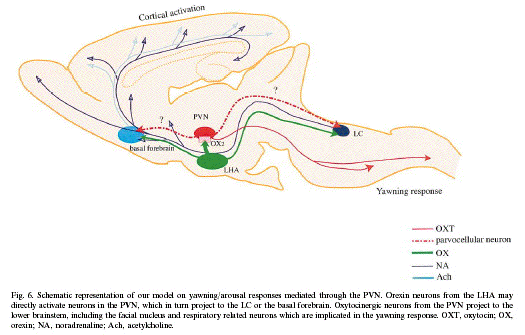
It is widely believed that activation of the
descending oxytocinergic system in the PVN is
one of the most important mechanisms for
mediating the induction of yawning (Argiolas et
al., 1987; Sato-Suzuki et al., 1987). However,
the PVN contains various neuropeptides besides
oxytocin, such as CRF, vasopressin, and
dynorphine (Swanson and Sawchenko, 1983). We
previously found, using c-Fos
immunohistochemistry in rats, that activation of
not only oxytocin neurons but also CRF neurons
in the PVN is involved in the yawning
accompanied by arousal response induced by PVN
stimulation (Kita et al., 2006a). The mechanism
by which CRF neurons produces yawning is
unclear, but nitric oxide (NO) produced within
CRF neurons may cause the yawning. NO is a
diffusible neurotransmitter/neuromodulator and
acts as a paracrine agent. Several studies have
demonstrated that co-localization of NO
synthesis with CRF-producing parvocellular in
the PVN (Harada et al., 1999; Siaud et al.,
1994; Torres et al., 1993). It is, thus,
possible that diffusible NO produced within CRF
neurons activates descending oxytocinergic
system in the PVN and its actions on
oxytocinergic neurons elicit yawning. This
possibility could be supported by previous
studies showing that NO could be involved in
induction of yawning via an activation of
oxytocinergic neurons projecting to the pons and
medulla oblongata (Argiolas et al., 1987;
Argiolas and Meus, 1998; Kita et al., 2006b;
Melis and Argiolas, 1997; SatoSuzuki et al.,
1998).
CRF neurons in the PVN generally play an
important role as the initiators of activation
of the hypothalamic-pituitary-adrenal axis (HPA
axis) during stress (Rivier and Plotsky, 1986;
Vale et al., 1981). In addition, CRF can serve
in extrahypophyseal brain regions as one of the
neurotransmitters most likely to be involved in
mediating the arousal response as well as
autonomic and behavioral responses to various
stressors (Dunn and Berridge, 1990; Heinrichs
and Koob, 2004; Menzaghi et al., 1993). Our
previous studies have shown that stress-like
stimulation could induce arousal response during
yawning (Kita et al., 2000; Seki et al., 2003).
In the present study, we showed that the CRF
antagonist despite its ability to reduce arousal
response measured by EC0G was unable to reduce
yawning, being consistent with our previous
study (Kita et al., 2008). It, thus, is possible
that CRF neurons in the PVN, which can be
activated by various stressors, primarily
mediate the arousal response during yawning
behavior, and that the oxytocinergic system in
the PVN might be responsible for behavioral
responses rather than the arousal response
during yawning behavior.
Although the CRF-containing pathway involved
in an arousal response during yawning evoked by
PVN stimulation is still
unknown, a series of animal studies has
suggested that CRF serves as an excitatory
neurotransmitter in the LC and the DRN (Curtis
et al., 2002; Jedema and Grace, 2004; Kirby et
al., 2000; Lowry et al., 2000; Schulz and
Lehnert, 1996; Smagin et al., 1995; Staub et
al., 2005), which are the regions within the
brainstem reticular formation responsible for
cortical activation. Several anatomical studies
have identified that the LC and DRN, which are
the major sources of NA and 5-HT in the central
nervous system respectively, receive CRF
afferents from the PVN (Curtis et al., 1997;
Dunn et al., 2004; Swanson et al., 1983;
Valentino et al., 1992), and project to various
areas of the cortex (Foote et al., 1983; Meloni
et al., 2008; Swanson and Hartman, 1975; Vertes,
1991). In addition, it is reported that CRF
terminal synaptically contact the
catecholaminergic dendrites of LC neurons and
serotonergic dendrites of DRN neurons, and that
the CRF receptor is expressed in the LC NA
neurons and DRN 5-HT neurons (Commons et al.,
2003; Roche et al., 2003; Van Bockstaele et al.,
1996, 1998; Waselus et al., 2009). Functional
studies have Q4 shown that local administration
of CRF into the LC or DRN affects neuronal
activity in the LC or DRN respectively (Curtis
et al., 2002; Jedema and Grace, 2004; Kirby et
al., 2000; Staub et al., 2005, 2006; Valentino
et al., 1983), whereas microinjection of the CRF
receptor antagonist greatly inhibits the
magnitude of the LC or DRN activation by stress
(Kirby et al., 2000; Roche et al., 2003; Schulz
and Lehnert, 1996; Smagin et al., 1995; Staub et
al., 2005). The present study showed that
microinjection of L-glutamate into the PVN
significantly increases c-Fos expression in both
LC NA neurons and DRN 5-HT neurons as well as
PVN CRF neurons, and induced the arousal
response during yawning. Furthermore, we also
found that icy administration of CRF antagonist
significantly suppressed increases in the
activity of LC NA neurons and DRN 5-HT neurons,
but not PVN CRF neurons, during yawning induced
by the PVN stimulation, and suppressed the
arousal response during yawning. It is, thus,
suggested that a neural pathway from PVN CRF
neurons to LC NA neurons and DRN 5-HT neurons is
involved in the arousal response during yawning.
However we cannot rule out the possibility that
the CRF antagonist (icy) may combine with CRF
receptor in several areas of the brain besides
LC and DRN, because we used icy administration
of the CRF antagonist in order to examine the
involvement of CRF in the arousal response
during yawning in the present study. It has been
reported that CRF receptor is widely distributes
in the several brain areas, such as cerebral
cortex, lateral septum nucleus, hippocampus,
amygdale, and hypothalamus as well as LC and DRN
(Aguilera et al., 2004; Chalmers et al., 1995;
Reul and Holsboer, 2002). However, our data
showed that icy administration of CRF antagonist
significantly suppressed increases in the
activity of LC and DRN neurons during yawning
induced by the PVN stimulation, as well as
significant attenuation of arousal response
during yawning. In addition, LC and DRN are
located in the periventricular area, in which
CRF antagonist delivered
intacerebroventricularly can be to exert its
dominant effects on (Bittencourt and Sawchenko,
2000; Pardridge, 1997). Therefore, it is
possible that icy administration of CRF receptor
antagonist could act on at least LC and DRN
neurons in the present study.
Several anatomical studies have indicated
multiple sources of CRF innervations to the LC
and DRN, including the nucleus
paragigantocellularis, Barrington's nucleus,
central nucleus of the amygdale, bed nucleus of
the stria terminals as well as the PVN (Curtis
et al., 2002; Dunn et al., 2004; Valentino et
al., 1992, 1996, 2010; Van Bockstaele et al.,
1998,1999). Although the PVN CRF neurons is one
of the most possible sources of CRF innervations
to the LC and DRN, which can mediate the arousal
response, we cannot rule out the possibility
that the other sources of CRF neurons projecting
to the LC and DRN may contribute to the arousal
response during yawning.
Furthermore, we cannot exclude the
involvement of oxytocincontaining pathway in the
arousal response during yawning behavior. It has
been shown that parvocellular oxytocin neurons
in the PVN are known to send descending axons to
the lower brain stem, including the LC (Schulz
and Lehnert, 1996; Sawchenko and Swanson, 1982).
In this study, pretreatment with CRF antagonist
could not completely suppress the arousal shift
in EC0G and the activation of LC NA neurons
during yawning induced by PVN stimulation.
Therefore, the oxytocin-containing pathway
projecting to the LC may also be involved in the
arousal response during yawning. This
possibility should be investigated further
through the use of oxytocin antagonists or
higher doses of CRF antagonists.
The present study showed that the c-Fos
active 5-HT neurons in the DRN were almost
completely suppressed by the CRF receptor
antagonist but NA neurons in the LC were not,
suggesting the functional difference of these
two neurotransmitter systems in controlling
arousal response during yawning. It has been
suggested that NA particularly is involved in
alarm and anxiety state (Elam et al., 1986;
Morilak et al., 2005; Prokopova, 2010; Ressler
and Nemeroff, 2000), and that 5-HT is involved
in anxiolitic and antidepressant behavior
(Graeff et al., 1996; Meloni et al., 2008;
Ressler and Nemeroff, 2000), though both NA and
5-HT are known to be stress-related
neurotransmitter as well as CRF neurons. Thus,
it is considered that the difference of neuronal
activity between NA and 5-HT neurons during
yawning can control the quality of arousal
response during yawning, for example, different
levels of emotional arousal, attention, or
cognition.
In summary, we observed that several
microinjections of L-glutamate into the PVN
produced frequent spontaneous yawning
accompanied by an arousal response, and the
arousal response is associated with significant
increases in the number ofc-Fos positive PVN CRF
neurons, LC NA neurons, and DRN 5-HT neurons.
However, pretreatment with CRF antagonist (icv)
significantly inhibited the activation of both
LC NA neurons and DRN 5-HT neurons, but not the
activation of PVN CRF neurons. It also
significantly suppressed the arousal response
during yawning. These results suggest that CRF
neurons in the PVN may mediate the arousal
response during yawning behavior through
activation of LC NA neurons and DRN 5HT neurons.
Further studies are needed to clarify the direct
pathway from PVN CRF neurons to LC NA neurons
and DRN 5-HT neurons responsible for arousal
response during yawning behavior.
-Collins GT, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior. J Pharmacol Exp Ther
2005;314(1):310-319.
-Collins GT, Newman
AH,Woods JH et al.Yawning and hypothermia in
rats: effects of dopamine D3 and D2 agonists and
antagonists. Psychopharmacology
(Berl).
2007;193(2):159-170
-Collins GT. et
al. Food restriction alters
pramipexole-induced yawning, hypothermia, and
locomotor activity in rats: Evidence for
sensitization of dopamine D2 receptor-mediated
effects. JEPT 2008;325:691-697
-Collins
GT et al. Narrowing in on compulsions:
dopamine receptor functions Exp Clin
Psychopharmacol 2008,16(4):498-502
-Collins
GT et al. Pro-erectile Effects of Dopamine
D2-like Agonists are Mediated by the D3 Receptor
in Rats and Mice JPEP 2009;329(1):210-217
-Collins GT,
Truong YN, et al. Behavioral sensitization
to cocaine in rats: evidence for temporal
differences in dopamine D(3) and D (2) receptor
sensitivity. Psychopharmacology (Berl).
2011;215(4):609-620.
-Hipolide DC;
Lobo LL; De Medeiros R; Neumann B; Tufik S
Treatment with dexamethasone alters yawning
behavior induced by cholinergic but not
dopaminergic agonist. Physiol Behav
1999;65(4-5):829-832
-Hipolide
DC, Tufik S Paradoxical sleep deprivation in
female rats alters drug-induced behaviors
Physiol Behav. 1995;57(6):1139-1143
-Kita I, Kubota N,
Yanagita S, Motoki C Intracerebroventricular
administration of corticotropin-releasing factor
antagonist attenuates arousal response
accompanied by yawning behavior in rats.
Neurosci.Letter 2008;433(3):205-208
-Kita I, Yoshida
Y, Nishino S. An activation of parvocellular
oxytocinergic neurons in the paraventricular
nucleus in oxytocin-induced yawning and penile
erection. Neurosci Res. 2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Beh Brain Res
2000;117(1-2):119-126
-Kubota N, Amemiya
S, Motoki C, Otsuka T, Nishijima T, Kita I.
Corticotropin-releasing factor antagonist
reduces activation of noradrenalin and serotonin
neurons in the locus coeruleus and dorsal raphe
in the arousal response accompanied by yawning
behavior in rats. Neurosci Res. 2012
-Moyaho
A, Valencia J Grooming and yawning trace
adjustment to unfamiliar environments in
laboratory Sprague-Dawley rats J Comp Psychol
2002;116(3):263-269
-Neumann BG,
Troncone LR, Braz S, Tufik S Modifications
on dopaminergic and cholinergic systems induced
by the water tank technique: analysis through
yawning behavior. Arch Int Pharmacodyn Ther
1990;308:32-38
-Sato-Suzuki I,
Kita I; Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5):2765-2775
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Tufik S et
al Effects of stress on drug induced yawning
Physiol Behav 1995;58(1):1881-1884