- Test
de susceptibilité à la contagion
du bâillement
-
- Contagious
yawning -
Réplication
du bâillement
bibliographie
complète
- Contagiosité
des comportements humains : la
réplication du bâillement peut-elle
nous éclairer
?
-
- Scipion
Dupleix 1626
 -
- Rambosson
J. Propagation à distance des affections
et des phénomènes nerveux
expressifs
- Mémoire
lu à l'Académie de médecine
le 8 juin 1880
-
-
- Les humains sont des êtres sociaux.
Une des fonctions primordiales de
l'encéphale est de nous donner la
capacité d'interagir de façon
optimale avec les autres. Le succès des
interactions sociales réside dans la
capacité à comprendre les autres
au niveau des actions motrices
(intentionnalité), des perceptions
émotionnelles, d'une intégration
cognitive mnésique et comparative
séparant le soi de l'autre (empathie,
altruisme), toutes capacités que le
psychologie anglo-saxonne a unifié sous
le concept de "The
theory of mind" (TOM). Il est connu, depuis
toujours, que le bâillement est
"contagieux"; l'éthologie parle de
réplication comportementale et, la
neurologie, depuis JM. Charcot,
d'échokinésie.
En quoi cette échokinésie
confère aux bâillements une forme
de communication sociale non verbale participant
de la TOM et de l'empathie ? (L'échokinésie
définie par Emile Littré).
Indiquons, enfin, que si le bâillement est
"contagieux", la pandiculation ne l'est pas, ce
pouquoi restant à élucider.
-
- La découverte des neurones miroirs
par Rizzolatti
et Gallese
(1996) ouvre à une explication
neurophysiologique de la TOM. La promotion des
compétences exploratrices de
l'environnement, les processus de prises de
décisions, notamment en cas d'urgence
face à un prédateur, la
préparation de l'action en
générale, répondent chez la
plupart des vertébrés à
l'activation de neurones miroirs dans les aires
corticales motrices. Ils s'activent lors de la
perception des mouvements et d'action de
conspécifiques indiquant que la
génération de l'action
intentionnelle et l'imagerie mentale de celle-ci
partagent les mêmes structures neuronales.
Ainsi, sur un trottoir parisien, l'approche du
piéton décelé par un pigeon
déclenche automatiquement l'envol de la
troupe dont la majorité n'a pas eu
perception du danger. Résultat de
réponses adaptatives
sélectionnées par
l'évolution, cet automatisme moteur
coopératif valorise la vie en groupe en
terme de sécurité face au
prédateurs.
-
- L'échokinésie du
bâillement ne répond pas à
ce mécanisme comme l'indique son
délai d'apparition et son inconstance. En
effet, n'est sensible à
l'échokinésie qu'un individu dans
un état mental peu stimulé
(transport en commun), alors qu'un travail
intellectuel soutenu le rend insensible.
-

- Mirror
neuron activity during contagious yawning-an
fMRI study
- Haker H, Kawohl W, Herwig U,
Rössler W.
- Brain Imaging
Behav. 2012
-
- Schürmann
et al.(2005) ont confirmé, par
exploration en IRM fonctionnelle, l'absence
d'activation des neurones miroirs de l'aire de
Broca
chez l'Homme (cortex frontal
postéro-inférieur de
l'hémisphère gauche), lors de
l'échokinésie du bâillement,
contrairement à l'observation gestuelle
pendant laquelle, ceux-ci s'activent tant dans
l'exécution que dans l'observation d'une
activité motrice (décodage de
l'intentionnalité). Ces
éléments éthologiques et
neurophysiologiques montrent que
l'échokinésie du bâillement
n'est pas une imitation motrice.
-
- Haker,
Rössler et al. (2012) ont, eux aussi,
utilisé l'imagerie par IRM fonctionnelle
pour visualiser l'activité neuronale
durant la contagion du bâillement. En
réponse à un bâillement, les
sujets montrent une activation unilaérale
de l'aire 9 de Brodman (BA 9), partie du gyrus
frontal inférieur, une région
où sont présents les neurones
miroirs.Par ces voies, deux individus peuvent
partager des états physiologiques et
émotionnels basés sur un pattern
moteur. C'est l'un des composants de l'empathie
(empathie motrice) qui sous-tend celui de
l'empathie cognitive. L'aire BA 9 est active au
cours de tâches requérant une
réfélexion. Ces résultats
soulignent la connection entre le système
des neurones miroirs et les fonctions cognitives
en rapport avec l'empathie.Ils concluent que la
contagion du bâillement est basée
sur le sytème neuronal actif lors de
l'empathie.
-
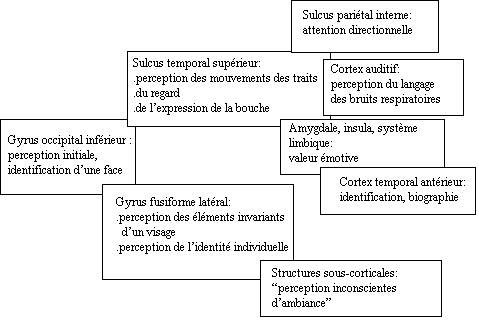
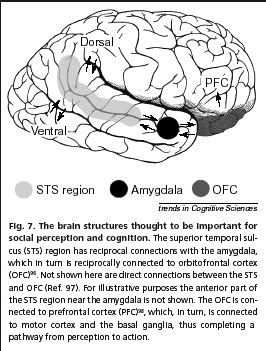
- La reconnaissance visuelle de
l'environnement met en jeu différents
circuits neuronaux distinguant les objets
inanimés des êtres vivants
(Puce,
2003). La reconnaissance des visages humains
répond à l'activation de neurones,
spécifiquement dédiés, au
niveau temporal. La région temporal
inférieur (IT) (Afraz,
2006; Leopold,
2006) permet une identification immédiate
d'un visage dans sa globalité, tant pour
l'identité que pour l'expression, en
possédant, apparemment, une
mémorisation autonome, non hippocampique.
Le sulcus
temporal supérieur (STS) s'active,
lui, spécifiquement lors de perception
des mouvements des yeux, de la bouche,
suggérant son implication dans la
perception visuelle des émotions.
Schürmann
et al. (2005) ont montré l'activation du
STS lors de l'échokinésie du
bâillement, de façon automatique et
involontaire, se transmettant vers la
région periamygdalienne gauche, le cortex
cingulaire postérieur et le
précuneus. Ces structures sont
associées à la discrimination des
émotions exprimées par la face
humaine et, notamment, dans
l'appréciation de la
véracité du ressenti
exprimé.
-
- Platek et
al. (2003;
2005)
ont trouvé une corrélation entre
activation des circuits neuronaux au-delà
du STS et traits de personnalités. "A
l'opposé de ceux qui ne sont pas
influencés par la vue d'un autre
bâilleur, les gens qui sont sensibles au
bâillement d'autrui reconnaissent plus
promptement l'image de leur propre visage,
identifie mieux l'état psychologique de
l'autre et exhibe moins de traits de
personnalité d'allure schizoïde. Ces
données suggèrent que la contagion
du bâillement a un lien avec la
capacité d'une part d'introspection et
d'autre part d'empathie". Les sujets
qualifiés d'empathiques, très
sensibles à l'échokinésie
du bâillement activent peu l'amygdale mais
le cortex cingulaire, alors que les sujets
qualifiés de schizoïdes, insensibles
à la réplication du
bâillement, activent l'amygdale (la peur
?) (Carr,
2003). L'étude de la neurophysiologie de
l'empathie retrouve une similarité des
zones activées (STS,
insula,péri-amygdale, cortex cingulaire,
cortex préfrontal droit). Ainsi, il
semble qu'alors que la compréhension de
l'intentionnalité (neurones miroirs
moteurs), le partage des émotions
(neurones miroirs de l'insula, amygdale et
cortex pariétal droit)
nécessitent, pour être
décodés, une activation neuronale
commune, action - perception,
simultanément à une inhibition
frontale (activation orbito-frontale)
inhibitrice de l'extériorisation motrice,
l'échokénsie du bâillement
n'a pas la capacité d'être
inhibée involontairement par absence
d'inhibition frontale. Par contre l'activation
temporo-pariétale droite autorise la
différenciation entre soi et l'autre,
identifiant, au niveau conscient, le rôle
déclencheur du bâillement de
l'autre (Decety,
2006). Le bâillement pourrait ainsi
illustrer la théorie simulationniste de
la TOM. En 2008, Nahab
et al. ont montré en IRMf que la zone
ventromédiale frontale s'activait au
cours de l'echokinésie du
bâillement, zone activée
également au cours des processus
empathiques et proposent de situer à ce
niveau la zone corticale à l'origine du
déclenchement du bâillement
"contagieux".
-
- Alors que le bâillement apparaît
universel chez les vertébrés, il
semble que seuls les primates soient capables
d'une échokinésie. Anderson
(2004) a montré que les chimpanzés
visionnant une vidéo de bâillements
de conspécifiques, bâillaient
à la suite, de façon similaire aux
humains dans une situation analogue. Bien que
l'existence d'une TOM chez le chimpanzé
soit toujours l'objet de controverses (Povinelli,
2003), l'existence d'une
échokinésie des bâillements
plaide en faveur d'une théorie de
différents niveaux de TOM,
peut-être secondairement à des
voies différentes de l'évolution
des capacités cognitives des
hominidés. La pathologie psychiatrique
humaine dissèque, elle aussi, de
façon comparable, la TOM (Blair,
2005).
-
- A.
Senju et al. (14) A. Senju et al. ont
projeté des videos clips de personnes
bâillant ou ouvrant simplement la bouche
à 49 enfants, agés de 7 ans, et
dont la moitié était des autistes.
La vision de bâillements
déclenchent plus du double de
bâillements chez les enfants sains que
chez les autistes. Regarder des visages ne
bâillant pas déclenche le
même nombre de bâillements dans les
deux catégories d'enfants. Cette
étude eoque donc un déficit
d'échokinésie des
bâillements chez les personnes autistes.
Or le déficit a décodé les
émotions des autres, ainsi que
l'empathie, sont des critères
diagnostiques de cette pathologie. Ces
données ont été
confirmées en
2010 par M. Helt et IM. Eigisti
-
- L'équipe de A. Senju
propose, en 2008, que sous certaines conditions
de lien de dépendance longtemps
établi, un chien pourrait bâiller
en voyant son maître bâiller. Les
mêmes seraient capables de comprendre
l'indication de la direction du regard
donnée par un doigt pointé vers
une cible ce qui montrerait une capacité
de TOM chez ces chiens.
-
- Empathy
in schizophrenia: impaired resonance Haker
H, Rössler W 2009 : dans ce travail, les
auteurs montrent la faible
réceptivité des
schizophrènes aux bâillements
d'autrui. Mais la prise d eneuroleptiques par
ceux-ci étant inhibitrice des
bâillements, la pertience de la
démontsration est réduite bien
théoriquement cohérente
-
- Anderson
(2003) a montré que l'enfant
n'était sensible à
l'échokinésie du bâillement
qu'au cours de la sixième année de
vie, c'est à dire après avoir
acquis la capacité de
réfléchir à ce que l'autre
pense, à attribuer des états
mentaux à autrui (Gergely,
2003). Un état de maturation cognitive
d'ordre fonctionnel est donc nécessaire
afin d'être sensible à
l'échokinésie du
bâillement.
-
- Les
psychopathes montrent une difficulté
à être sensible aux
bâillements des autres
-
- En résumé, un lien
phénoménologique apparaît
entre la capacité à attribuer un
état mental à autrui (TOM), base
de l'empathie et l'échokinésie du
bâillement. A côté de la
hiérarchie neuro-anatomique
séparant la TOM en niveaux
sensorimoteurs, émotionnels et cognitifs
((Singer, 2006), l'échokinésie du
bâillement autorise une dissociation de la
TOM, par son ontogenèse et sa
phylogenèse, en différents niveaux
développementaux, sous-tendus par une
activation différenciée de
circuits neuronaux spécifiques.
L'échokinésie du bâillement
aurait conféré un avantage
sélectif, en permettant une
synchronisation efficace des niveaux de
vigilance entre les membres d'un groupe social.
Elle participerait d'une forme d'empathie
instinctive involontaire, qualifiable de
rudimentaire, probablement apparue tardivement
au cours de l'évolution des
hominidés.
 -
- Contagious
yawning: the role of self-awareness and mental
state attribution Platek SM, Critton SR,
et al
- Yearning
to yawn: the neural basis of contagious
yawning Schurmann M, Hari R et
al
- Contagious
yawning and the brain Platek S,
Mohamed F, Gallup G
- Contagious
yawning in chimpanzees
Anderson JR
- Video-induced
yawning in stumptail macaques Paukner A,
Anderson JR
- Evidence
for contagious behaviors in budgerigars
(Melopsittacus undulatus): An observational
study of yawning and stretching. Miller ML,
Gallup AC et al.
-
- A
clinical and psychological study of
echo-reactions Srengel Ed, Vienna MD, Edin
LR 1947
-
- Empathy
in schizophrenia: impaired resonance Haker
H, Rössler W
-
- Yawning:
unsuspected avenue for a better understanding of
arousal and
interoception
- Le
bâillement: son histoire intérieur
ou neurophysiologie du bâillement revue
- au
travers de son ontogenèse et de sa
phylogenèse
O. Walusinski
-
 -
- 1 -
Rizzolatti G, Fadiga L, Gallese V, et al.
Premotor cortex and the recognition of motor
actions. Brain Res Cogn Brain Res.
1996;3:131-141.
-
- 2 -
Schürmann M, Hesse MD, Stephan KE, et
al. Yearning to yawn: the neural basis of
contagious yawning. Neuroimage.
2005;24:1260-1264.
-
- 3 - Puce
A, Perrett D. Electrophysiology and brain
imaging of biological motion. Philos Trans R Soc
Lond B Biol Sci. 2003;358:435-445.
-
- 4 - Afraz
SR, Kiani R, Esteky H. Microstimulation of
inferotemporal cortex influences face
categorization. Nature. 2006;442:692-695.
-
- 5 - Platek
SM, Mohamed FB, Gallup GG Jr. Contagious
yawning and the brain. Brain Res Cogn Brain Res.
2005;23:448-452.
-
- 6 - Platek
SM, Critton SR, Myers TE, Gallup GG.
Contagious yawning: the role of self-awareness
and mental state attribution. Brain Res Cogn
Brain Res. 2003;17(2):223-237.
-
- 7 -
Carr L, Iacoboni M, Dubeau MC, et al. Neural
mechanisms of empathy in humans: a relay from
neural systems for imitation to limbic areas.
Proc Natl Acad Sci USA. 2003;100:5497-5502.
-
- 8 - Decety
J, Grezes J. The power of simulation:
imagining one's own and other's behavior. Brain
Res. 2006;1079:4-14.
-
- 9 - Anderson
JR, Myowa-Yamakoshi M, Matsuzawa T.
Contagious yawning in chimpanzees. Proc Biol
Sci. 2004;271 Suppl 6:S468-470.
-
- 10 - Povinelli
DJ, Vonk J. Chimpanzee minds: suspiciously
human? Trends Cogn Sci. 2003;7:157-160.
-
- 11 - Blair
RJ. Responding to the emotions of others:
dissociating forms of empathy through the study
of typical and psychiatric populations.
Conscious Cogn. 2005;14:698-718.
-
- 12 - Anderson
JR, Meno P. Psychological influences on
yawning in children. Current Psychology Letters
Behaviour, Brain, & Cognition.
2003;2:390.
-
- 13 - Singer
T. The neuronal basis and ontogeny of
empathy and mind reading: Review of literature
and implications for future research. Neurosci
Biobehav Rev. 2006;30:855-863.
-
- 14 - Senju
A, Maeda M, Kikuchi Y et al. Absence of
contagious yawning in children with autism
spectrum disorder. Biology letters
2007;3:706-708.
-
- 15 -Joly-Mascheroni
RM, Senju A, Sheperd AJ Dogs catch human
yawns Biology letters Animal Behaviour
2008;4(5):446-448.
-
- 16 -Nahab
FB, Hattori N, Saad ZS, Hallett M Contagious
yawning and the frontal lobe: An fMRI study
Human Brain Mapping 2008 in press
-
 -
- Response
properties of neurons in
temporal cortical visual areas of infant
monkeys Rodman HR
-
-
Un précurseur des recherches actuelles...
oublié ! Yawning,
Yielding, and Yearning to
Yawn
- Paper presented at the
meeting of the Midwest Psychological
Assiociation Chicago, May
1974
- Cialdini
RB, McPeek RW
-
- Je suis heureux de pouvoir vous
présenter un document inédit
jamais publié. Ce travail a
été présenté
oralement, par Robert
Cialdini, en 1974, à l'association de
Pyschologie du Midwest.
-
- "Comme manger, bâiller est un
évènement quotidien qui peut aussi
bien être déclenché par la
fatigue (cause interne) que par la vision d'un
autre bâilleur (cause externe).
L'étude a été bâtie
pour:
- démontrer l'existence d'un
bâillement de stimulation externe,
à fonction sociale, et
- étudier la variabilité de la
susceptibilité individuelle à
recevoir cette contagion du
bâillement.
- .............
- Les résultats suggèrent que
des facteurs classiquement plus importants pour
leur influence sur la vie sociale sont mis
à l'oeuvre dans la contagion du
bâillement, eux-mêmes étant
contagieux. En particulier, l'usage de batteries
de tests jugeant de la sensibilité
"persuasive" indique que ceux qui sont le plus
facilement persuadés, sont
également plus susceptibles d'être
sensibles au bâillement des autres!
-
- On peut en déduire que la
crédibilité du pourvoyeur de
bâillements affecte le degré de
contagion.
- Capuchin
monkeys display affiliation toward humans who
imitate them.
-
- Paukner A, Suomi SJ, Visalberghi E, Ferrari
PF.
-
- Laboratory of Comparative
Ethology, Eunice Kennedy Shriver National
Institute of Child Health and Human Development,
National Institutes of Health Animal
Center
-
- Science.
2009;325(5942):880-883.
-
- Abstract
-
- During social interactions, humans often
unconsciously and unintentionally imitate the
behaviors of others, which increases rapport,
liking, and empathy between interaction
partners. This effect is thought to be an
evolutionary adaptation that facilitates group
living and may be shared with other primate
species. Here, we show that capuchin monkeys, a
highly social primate species, prefer human
imitators over non-imitators in a variety of
ways: The monkeys look longer at imitators,
spend more time in proximity to imitators, and
choose to interact more frequently with
imitators in a token exchange task. These
results demonstrate that imitation can promote
affiliation in nonhuman primates. Behavior
matching that leads to prosocial behaviors
toward others may have been one of the
mechanisms at the basis of altruistic behavioral
tendencies in capuchins and in other primates,
including humans.
-
- -Arnott
SR et al. An investigation of auditory
contagious yawning Cognitive, Affective,
Behavioral Neurosci 2009;9(3):335-342
- -Anderson
JR et al Contagious yawning in chimpanzees
The Royal Society Biology Proc R Soc Lond B Biol
Sci 2004; 271 Suppl 6; S468-470
- -Anderson
JR et al Psychological influences on yawning
in children Current Psychology Letters
Behaviour, Brain, Cognition 2003;2:11
- -Anderson
JR, Matsuzawa T. Yawning: An Opening into
Empathy? Cognitive Development in Chimpanzees. T
Matsuzawa, M Tomonaga, M Tanaka Editors.
Springer 2006
- -Caswell
TA, The effects of status on yawning
behavior. Thesis 1991
- -Cialdini
RB, McPeek RW. Yawning, Yielding, and
Yearning to Yawn 1974 (non published)
- -Campbell
MW et al. Do chimpanzees yawn
contagiously in response
to 3d computer animations? 2008
- -Campbell
MW et al. Computer animations stimulate
contagious yawning in
chimpanzees Proceed Royal Soc Biol
2009:276(1676):4255-4259
- -Campbell
MW, de Waal FB. Ingroup-Outgroup Bias in
Contagious Yawning by Chimpanzees Supports Link
to Empathy. Plos One. 2011;6(4):1-4
- -Cooper
NR, Puzzo I, Pawley A Contagious yawning:
the mirror neuron system may be a candidate
physiological mechanism Medical Hyportheses
2008;71(6):975-976
- -Cooper
NR, Puzzo I, et al. Bridging a yawning
chasm: EEG investigations into the debate
concerning the role of the human mirror neuron
system in contagious yawning.Cogn Affect Behav
Neurosci. 2011
- -Derouesné
Ch. Théorie de l'esprit, empathie et ..
bâillement 2005
- -Dijksterhuis
A, Bargh JA The perception-behavior
expressway:automatic effects of social
perception on social behavior Advances in
Experimental Social Psychology
2001;33:1-40.
- -Estow,
S Jamieson JP, Yates JR Self-monitoring and
mimicry of positive and negative social
behaviors Journal of Research in Personality
2007;41(2):425-433
- -Gallagher
HL, Frith CD Functional imaging of theory of
mind Trends in Cognitive Scie.
2003;7(2):77-83
- -Giganti
F, Ziello ME Contagious and spontaneous
yawning in autistic and typically developing
children CPL 2009
- -Giganti
F, Zilli I. The daily time course of
contagious and spontaneous yawning among humans.
J Ethol 2011;29(2):215-216
- -Haker
H, Kawohl W, Herwig U, Rössler W.
Mirror neuron activity during contagious
yawning-an fMRI study. Brain Imaging Behav.
2012
- -Harr
AL, Gilbert VR Do dogs show contagious
yawning ? Anim Cogn. 2009;12(6):833-837
- -Helt
MS, Eigsti IM, Snyder PJ, Fein DA.
Contagious yawning in autistic and typical
development. Child Dev.
2010;81(5):1620-1631
- -Iacoboni
M, G Rizzolatti Cortical mechanisms of human
imitation Science 24/12/99; 286
- -Joly-Mascheroni
RM, Senju A, Sheperd AJ Dogs catch human
yawns Biology letters Animal Behaviour
2008;4(5):446-448
- -Lindsay
SR Coping with fear and stress: licking and
yawning. Handbook of applied dog behavior and
training 2000
- -Madsen
EA, Persson T. Contagious yawning in
domestic dog puppies (Canis lupus familiaris):
the effect of ontogeny and emotional closeness
on low-level imitation in dogs. Anim Cogn.
2012
- -Massen
JJ, Vermunt DA, Sterck EH. Male Yawning Is
More Contagious than Female Yawning among
Chimpanzees (Pan troglodytes). PLoS One.
2012;7(7):e40697.
- -Millen
A, Anderson JR Neither infants nor toddlers
catch yawns from their mothers. Biol Letters
2011;7(3):440-442
- -Miller
ML, Gallup AC, Vogel AR, Vicario SM, Clark
AB. Evidence for contagious behaviors in
budgerigars (Melopsittacus undulatus): An
observational study of yawning and stretching.
Behav Processes. 2012;89(3):264-270
- -Nahab
FB, Hattori N, Saad ZS, Hallett M Contagious
yawning and the frontal lobe: An fMRI study
Human Brain Mapping 2009;30:1744-1751
- -Norscia
I, Palagi E. Yawn Contagion and Empathy in
Homo sapiens. PLoS ONE. 2011;6(12): e28472
- -O'Hara
SJ, Reeve AV A test of the yawning contagion
and emotional connectedness hypothesis in dogs,
Canis familiaris. Animal Behaviour
2011;81:335-340
- -Palagi
E, Leone A, Mancini G, Ferrari PF.
Contagious yawning in gelada baboons as a
possible expression of empathy. Proc Natl Acad
Sci USA. 2009;106(46):19262-19567
- -Paukner
A, Anderson JR
Video-induced yawning in stumptail
macaques (Macaca
arctoides) Biol Lett 2006;2(1):36-38
- -Perkins
JR Teaching Dogs to Yawn, Sneeze, and
Implications for Preparedness Theory and
Observational Learning.
In: Kusonose, Ryo and Sato, Shusuke 39th
Congress of the International Society for
Applied Ethology, Kanagawa, Japan. 20-24 August,
2005
- -Platek
SM, SR Critton, et al Contagious yawning:
the role of self-awareness and mental state
attribution Cogn Brain Res
2003;17(2):223-227
- -Platek
S et al. Neural correlates of self-face
recognition 2008 Brain Res;1232:173-184
- -Platek
S, Mohamed F, Gallup G Contagious yawning
and the brain Cognitive Brain Research,
2005;23:448-452
- -Schurmann
M, Hari R et al Yearning to yawn: the neural
basis of contagious yawning NeuroImage
2005;24(4):1260-1264
- -Senju
A, Maeda M, Kikuchi Y, Hasegawa T, Tojo Y,
Osanai H Absence of contagious yawning in
children with autism spectrum disorder. Biology
Letters 2007;3:706-708
- -Senju
A, Kikuchi Y, Akechi H et al. Does eye
contact induce contagious yawning in children
with autism spectrum disorder? J of Autism and
Developmental Disorders.
2009;39(11):1598-1602
- -Silva
K, Bessa J, de Sousa L. Auditory contagious
yawning in domestic dogs (Canis familiaris):
first evidence for social modulation. Anim Cogn.
2012.
- -Yoon
JMD, Tennie C Contagious yawning: a
reflection of empathy, mimicry, or contagion.
Anim Behav 2010;79:e1-e3
- -Walusinski O
Echokinetic yawning, theory of mind, and
empathy
- -de
Waal F. l'âge de l'empathie. 2010
- -Wilkinson
A et al. No evidence of contagious yawning
in the red-footed tortoise. Current Zoology
2011;57(4):477-484
|


