Effects of single
and repeated treatment with antidepressants on
apomorphine-induced yawning in the rat: the
implication of alpha-1 adrenergic mechanisms in
the D-2 receptor function
A. Delini-Stula, C. Hunn
Research Laboralories,
Pharmaceuticals Division, Ciba_Geigy Basle.
Switzerland
Yawning behaviour, induced by low doses of
apomorphine, is considered to be a functional
expression of a selective stimulation of
dopamine (DA)-receptors, which in contrast to
the earlier assumptions (Yamada
and Furukawa 1980, Stahle
and Ungerstedt 1984), appear to be localised at
the postsynaptic neuronal membrane (Serra et al.
1986, Morelli
et al. 1986; Stahle and Ungerstedt 1989). There
is convincing evidence demonstrating that
yawning is dominantly mediated by D-2 type
receptors (Protais
et al. 1983, Holm and Ôgren 1985), but a
contributory role of D-1 receptors bas also been
suggested by more recent findings (Stahle and
Ungerstedt 1989). Recently Mogilnicka
et al. (1986) reported that desipramine,
imipramine and some other antidepressants induce
yawning behavior in rats. This effect was
selectively antagonized by DA-receptor blocking
drugs. Therefore, it appeared to be of
doparninergic origin and unrelated to NA- or
5-HT inhibiting properties of the investigated
drugs.
Several antidepressants and electroconvulsive
treatment were, however, found to reverse the
apomorphine induced hypomotility in rats, which
is also of dopaminergic origin and thought to
result from the stimulation of the same
receptors as involved in the mediation of
yawning (Serra et al. 1979, 1981).
Antidepressants lack direct effects on DA
receptors and the mechanisms of these observed
effects have, up to now, remained unexplained.
The aim of the present study was to test the
assumption that, by virtue of their NA-enhancing
properties, antidepressants nevertheless
indirectly modulate D-2 receptors activated by
low doses of apomorphine.
Yawning behaviour was selected as a suitable
model for this purpose. Even though this might
be an overinterpretation, there are reasons to
assume that yawning reflects the vigilance state
of the animals.
The effects of chiral drugs, the (+)- and
(-)-enantiomers of oxaprotiline and of highly
selective 5-HTuptake inhibitors citalopram and
ifoxetine (Waldmeier et al. 1986) were compared
to standard antidepressants (desipramine,
imipramine, maprotiline), which, however,
display different NA- and 5-HT-uptake inhibiting
profiles. Of the two chiral compounds the ( +
)-oxaprotiline is a stereospecific and highly
selective inhibitor of NA-uptake, whereas the (
-)-oxaprotiline (levoprotiline) lacks any
influence on NA or 5-HT-uptake (Waldmeier et al.
1982). In distinct difference to
(+)-oxaprotiline, levoprotiline also fails to
down-regulate beta-adrenergic receptors or
NA-induced depression of cortical neurons after
chronic administration (Delini-Stula et al.
1983). In addition to these drugs, adrafinil
(CRL 40 028), a central acting alpha-1 receptor
agonist (Rambert et al. 1986), and prazosin, a
selective alpha-1 receptor antagonists, were
also tested. [...]
Discussion
The results of this study indicate that
apomorphine-induced yawning is consistently
modified only by those antidepressants which
primarily affect noradrenergic
transmission. However, none of these drugs
showed an intrinsic enhancement of yawning. This
is in contrast to the observation of Mogilnicka
et al. (1986), who found that desipramine and,
to a lesser extent, imipramine induced yawning
in rats. The reason for this discrepancy is
unclear, but does not seem to be due to
different doses of the drugs, since they were
about the same in her and our study. Selective
5-HT-uptake inhibitors citalopram and ifoxetine
failed to show consistent or significant effects
in the interaction with apomorphine, either
after acute or repeated administration. They
showed, however, some intrinsic enhancing action
on yawning. It is worth mentioning that
ifoxetine is a highly selective, but atypical,
inhibitor of 5-HT-uptake in that it exerts its
action only centrally and is practically devoid
of any direct antagonistic action on brain
receptors (Waldmeier et al. 1986).
The importance of NA component in the
inhibition of yawning is strengthened by the
stereoselectivity of action of the
(+)-enantiomer of oxaprotiline. Also, apart from
being a potent and selective, NA-uptake
inhibitor (+)-oxaprotiline is pharmacologically
a rather specific acting drug. It shows weak or
no affinity for beta-adrenergic, dopaminergic,
5-HT-2 and, most importantly, cholinergic
receptors (for references see Delini-Stula 1986
and unpublished). A cholinergic link in the
initiation of yawning has been demonstrated by
Yamada and Furukawa (1980) and anticholinergic
properties of a drug might have been implicated
in the suppression of this behaviour. Complete
lack of an effect of the (-)enantiomer
levoprotiline further indicates that H-1
receptor blockade (which both compounds have in
common) is of no relevance to yawning. Further,
it does not seem that any direct enhancing
effect on postsynaptic D-2-receptors is
implicated in this inhibitory action. As several
recent studies have shown, both (+) and
(-)enantiomers have:
- a) a common property to increase
amphetamine- and apomorphine-induced
stereotypies after single doses of 10 and 30
mg/kg IP with (-) form being even more active
than the (+)-one (Delini-Stula and Mogilnicka
1988),
- b) after chronic administration both drugs
equally enhance behavioral stimulation induced
by d-amphetamine and dopamine (Maj and Wedzony
1988)
- c) enhanced behavioural stimulation after
chronic treatment with (-)-enantiomer could be
antagonized by sulpirid (Delini-Stula et al.
1988).
The absence of acute effects of imipramine
and maprotiline seems, however, to contradict
the implication of noradrenergic mechanisms in
the modulation of yawing.
Both drugs are also inhibitors of NA uptake,
and maprotiline even a selective one. However,
both drugs directly act upon various other
receptor and transmitter systems as well.
Particularly, they have rather prominent alpha-1
receptor blocking properties. If our assumption
is true, than it is plausible to explain the
lack of their acute effects on yawning, among
others, by the counterbalancing alpha 1 receptor
blockade. However, after subchronic treatment
with these drugs yawning was markedly and
significantly inhibited. This is compatible with
the reported increased functional sensitivity of
alpha-1 receptors observed after repeated
administration of antidepressants (Mogilnicka et
al. 1987). Of interest, in this respect, is that
no change in specific 3H-prazosin binding was
observed after subchronic treatment with
antidepressants, but some findings suggest that
increased responsiveness of alpha-1 receptors
relates to their increased preference for the
agonists (Menkes et al. 1983). In the case of
imipramine, a part of its subchronic effect
could be due to the increased accumulation of
desipramine to which it is metabolized under in
vivo conditions. The implication of
noradrenergic mechanisms and, specifically,
alpha-1 receptors in the modulation of yawning
is, however, more directly supported by
clear-cut opposite effects of adrafinil and
prazosin.
The pharmacological profile of adrafinil, the
2/(diphenylmethyl)-sulfinyl/N-hydroxyacetamide,
indicates that this compound possesses selective
central alpha-l adrenergic agonistic properties.
The central action of adrafinil is evidenced by
the fact that behavioural activation is not
paralleled by signs of peripheral sympathetic
overstimulation. Selective alpha-1 adrenergic
properties are, on the other hand, supported by
the fact that behavioural hyperactivity, without
features of stereotypies, could be antagonized
by alpha-adrenergic antagonists such as
phenoxybenzamine, prazosin and yohimbine, but it
is unaffected by propranolol or pimozid (Duteil
et al. 1979; cf. Weetman 1980; Rambert et al.
1986). Our own investigations corroborate these
rindings. We also found that adrafinil in doses
of 30-120 mg/kg increases organized exploratory
and locomotor activity of rats in the open
field, enhances the plus-maze behaviour of mice
and reduces the acquired immobility in rats.
This behavioural activation, as reported by
other authors, was qualitatively different from
that induced by dopamineagonistic drugs.
Particularly, it was lacking stereotyped
features (Delini-Stula and van Riezen,
unpublished).
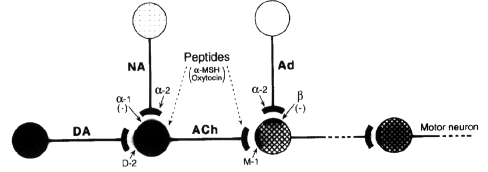
Altogether, the results of our study indicate
that, by virtue of their noradrenergic-enhancing
properties, antidepressants inhibit yawning
behaviour induced by apomorphine. The findings
also offer indirect evidence that the function
of D-2 dopamine receptors, activated by low
doses of apomorphine, is under modulatory
control of alpha-1 adrenoceptors. The
attractivness of this assumption consists in
that is suggests a link between the alpha-1
adrenergic and D-2 receptors and their mutual
involvement in the control of vigilance.
Certainly, this assumption needs further and
more direct supportive evidence.