-
- Test
de susceptibilité à la
contagion du
bâillement
- Contagious
yawning -
Réplication
du
bâillement
bibliographie
complète
- Contagiosité
des comportements humains : la
réplication du bâillement
peut-elle nous éclairer
?
-
- Scipion
Dupleix 1626
 -
- Robert
Whytt (1714-1766)
- "There is still a more
wonderful sympathy between the nervous system
of different persons"
- Observations on
the nature, causes and cure of those
disorders which have been commonly called
nervous, hypochondriac or hysteric, (1767)
-
- Shakespeare: «Nous ne
connaissons les autres que par
nous-mêmes et nous vibrons à
l'unisson pour les mieux
connaître».
-
- Yawning:
unsuspected avenue for a better understanding
of arousal and
interoception
- Echokinetic
yawning, theory of mind, and
empathy
- Rambosson
J. Propagation à distance des
affections et des phénomènes
nerveux
expressifs.
- Mémoire
lu à l'Académie de
médecine le 8 juin
1880
-
-
- Un célèbre dicton populaire
prédit qu'"un bon bâilleur en
fait bâiller sept". Il est coutumier de
parler de contagion. Ce terme est
adapté aux maladies infectieuses
quand, par contact direct ou indirect, un
homme sain devient malade en raison de la
transmission d'un virus ou d'un microbe. La
réplication d'un comportement ne
sous-entend aucune transmission d'un agent
quelconque. Il semble donc plus précis
de parler d'imitation comportementale, de
synchronisation d'actions. C'est pouquoi nous
userons du terme de réplication pour
cet exposé.
-
- L'étude éthologique des
primates (10) révèle
qu'à certains moments, un groupe
entier se met à bâiller
ensemble, sans qu'un individu ne puisse
percevoir l'autre de quelque manière
que ce soit, ni visuellement, ni
auditivement, ni olfactivement. Il ne peut
donc pas être considéré
qu'un tel comportement soit comparable
à la réplication interhumaine,
mais est lié, par exemple, à la
reprise d'activité de façon
synchrone, en raison des rythmes circadiens
repos - activité. Evidemment, chez
l'homme dans certains cas, ces deux variantes
peuvent se confondre.
-
- Cette réplication de
bâillement est initiée
involontairement. Aussi bien le
bâilleur n'éprouve aucun
désir de faire bâiller, que le
spectateur - receveur de la
réplication n'a conscience d'un
désir de bâiller. Le
bâillement de ce dernier est, lui
aussi, initié de façon
totalement involontaire, mais seulement et
seulement si son niveau de vigilance
l'autorise. En effet, l'implication dans une
tâche intellectuelle soutenue (c'est
à dire avec une concentration
élévée ou un niveau de
vigilance optimum) ne permettra pas le
déclenchement du bâillement.
Ce point est fondamental pour
l'interprètation éthologique
humaine du rôle de synchronisation des
états de vigilance entre deux
individus soumis à la transmission du
bâillement.
-
- Comment se déclenche cette
réplication ? La vue est un puissant
stimulant. Provine
(34,35) montre que 55% des spectateurs d'une
vidéo montrant 30 bâillements
successifs vont bâiller dans les cinq
minutes. Le temps de latence varie de
quelques secondes à cinq minutes.
Temps de latence et durée de la
visualisation n'ont pas permis
d'établir une règle
précise, un type de synchronisation
spécifique. Des sourires
répétés, en utilisant la
même technique que la vidéo des
bâillements, n'engendrent que quelques
rares sourires, indiquant ainsi la
spécificité de la
réplication de ce comportement de
bâillement.
-
- R.
Provine (36,37,38) a également
vérifié que le visage du
bâilleur n'a aucunement besoin
d'être dans un axe visuel précis
par rapport à celui qui subit la
réplication. Face à face,
à 90°, 180°, 270° l'un
de l'autre, la réplication a lieu.
L'existence d'une susceptibilité
à la réplication des aveugles
confirme que la vue n'est pas le seul
déclencheur stimulant. La vue d'une
partie seulement du visage, comme la bouche
largement ouverte, ne déclenche pas la
réplication. Il existe donc la
nécessité d'une perception
multimodale de toute la configuration du
visage et des temps respiratoires audibles
avec une dynamique coordonnée pour que
la réplication se réalise.
Alors que la présentation de
vidéo de bâillements
répétés déclenche
la réplication, la visualisation de
dessins animés, ne comportant qu'une
simplification des traits du visage et de la
mimique mobile du bâilleur ne
permettent pas la réplication.
D'autres mécanismes que la vue et
l'audition peuvent déclencher des
bâillements: la suggestion, la
lecture d'un texte sur le
bâillement (vous en ce moment ?),
l'évocation par la pensée
peuvent ainsi déclencher un
bâillement.
 - La réplication du bâillement
semble se situer à un niveau "basique"
car elle est indépendante de la
connaissance préalable du
déclencheur, indépendante de
caractères raciaux, éducatifs,
socio-culturels témoignant de
l'absence d'intervention mnésique. Il
n'y a pas besoin de caractérisation
explicite de l'autre pour subir la
réplication. Il faut être
à un niveau de vigilance
intermédiaire entre somnolence et
concentration soutenue, percevoir l'autre de
façon inconsciente mais être
capable de percevoir une chronologie
rigoureuse de la cinématique du
bâillement par ces composantes
visuelles et ou sonores de l'ouverture de
bouche, des modifications spécifiques
des autres traits faciaux qui s'y associent,
des mouvements et bruits de la ventilation
reflets de l'inspiration ample et
prolongée, de l'acmé, et
à un moindre degré de
l'expiration (35,36,37).
-
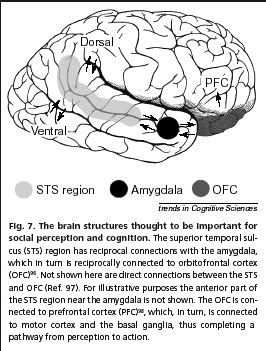
- Quels en sont les substrats
neurophysiologiques (20,21)? L'expertise
visuelle de reconnaissance faciale (2,11,15)
fait intervenir une grande
variété de processus,
réponses spécifiques à
un besoin social: reconnaissance anatomique
d'un visage, du sexe, de l'age, de ses
composants fixes et mobiles avec
critères de vitesse, de coordination,
d'harmonie ou de dysharmonie, des expressions
qu'ils véhiculent etc... L'examen des
déficits corticaux comme la
cécité cérébrale
(18), la prosopagnosie, par exemple, montre
qu'à côté de la
perception consciente existe un grand nombre
d'informations "d'ambiance" de la
scène regardée qui ne
franchissent pas le seuil de la conscience
mais participent implicitement et
obligatoirement à la perception
d'ensemble (6).
-
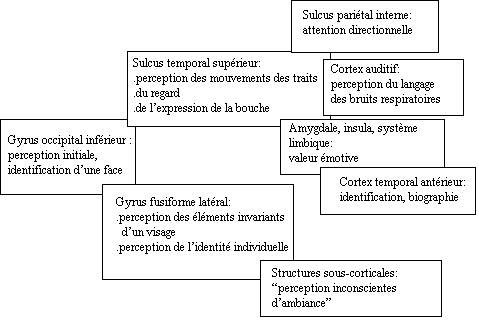
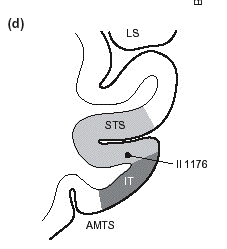
- La perception fait intervenir
différentes régions anatomiques
des deux hémisphères :
- - structures du cortex visuel, par trois
parties des régions
occipito-temporales: gyrus occipital
inférieur, gyrus fusiforme
latéral, sulcus
temporal supérieur. (voir
schéma ci-dessous et bas de page)
- - structures du cortex auditif : noyaux
cochléaires du tronc
cérébral, noyaux olivaires
supérieurs, lemnis latéral vers
le colliculus inférieur, corps
genouillé médian du thalamus,
cortex temporal.
- Les structures sous corticales
gèrent ces perceptions par des boucles
amygdale-thalamus-insula -système
limbique, avec des projections vers le tronc
cérébral. Les réactions
émotionnelles inconscientes sont
supportées par ces modalités
baptisées modèle perceptuel et
moteur des
émotions (19).
 - Une fois la perception acquise, le
déclenchement moteur du réflexe
de bâillement est involontaire et
résulte de mise en action de boucles
motrices sous-corticales noyaux gris - tronc
cérébral (16). En
parallèle, existe une extraction
consciente du déroulement du
phénomène, de son stimulus et
de sa valence contextuelle par les voies de
l'interroception
(5) permettant une perception
hédoniste consciente. Les voies tronc
cérébral - noyau
ventro-médian du thalamus - cortex
insulaire - cortex orbitofrontal sont alors
mises en jeu (11,20).
-
- D'autres comportements se
révèlent contagieux : le
rire (le rire et le cri pathologiques
sont associés à des
dysfonctions des voies de communications
réciproques - cortex cingulaire
préfrontal antérieur - cervelet
), les émotions (19) avec par exemple
la peur, génératrice de fuites
collectives; d'autres structures corticales
sont en jeu.
-
- Après avoir distingué les
réactions programmées, communes
à tout le monde animal et les
réactions émotionnelles
élaborées, propres aux
primates, il apparaît que la
réplication du bâillement peut
participer des deux types de comportements
(25).
-
- Les réactions
programmées sont retrouvées
chez tous les mammifères. Elles
apparaissent nécessaires à la
survie individuelle ou de groupe. Elles sont
contagieuses par communication non verbale.
Il s'agit d'un processus cognitif de
communication directe, immédiat non
conscient, engendrant des schémas
moteurs innés ou acquis telle la fuite
ou l'évitement. Baldwin, en 1894 (3),
a baptisé mimétisme ce type de
comportement. Le bâillement contagieux
ne peut correspondre que pour une part
à ce niveau comportemental
archaïque. Son automatisme l'en
rapproche. Mais son caractère
aléatoire, sa latence
éventuelle sont bien
différents.
-
- Une émotion exprimée
par un congénère
nécessite un traitement analytique de
l'information afin d'être
décryptée. Base de la
cognition sociale (1,28), l'expression
faciale des émotions sous-tend des
processus cognitifs élaborés et
flexibles. Leurs mises en oeuvre et la
réponse adaptée
nécessitent un temps de
réaction nettement plus important, mal
adapté à un réflexe
immédiat de survie. Seuls les primates
humains manifestent la capacité de
percevoir les autres comme des agents
intentionnels, avec capacité
d'identification. Cette capacité
à penser l'autre, à comprendre
le raisonnement d'autrui et ses désirs
sont cruciaux pour l'acquisition de
compétences à une vie sociale
(Theory of
mind) (12,42). C'est la base de
l'empathie réfléchie (7) :
comprendre le ressentiment de l'autre,
ressentir soi-même ce que l'autre
ressent. La réplication du
bâillement s'apparente au
décryptage d'une émotion, d'un
état de vigilance d'autrui à un
niveau automatique non conscient permettant
une synchronisation d'état de
vigilance entre individus, qu'on pourrait
qualifier d'empathie
instinctive involontaire (43). Cette
capacité à partager des
émotions avec autrui
(résonnance avec des affects
inconscients) repose sur une communication
implicite, façonnée au cours de
l'évolution, dont les
mécanismes neurobiologiques commencent
à s'éclairer
(1,2).( Yearning to
yawn: the neural basis of contagious
yawning MD Hesse, R Hari)
-
- «L'analyse des substrats
neurologiques des comportements montre un
caractère modulaire permettant une
explication évolutive par
accroissement graduel des composants. Des
structures neuronales spécifiques sous
tendent ce traitement de l'information
émotionnelle: l'amygdale, le cortex
occipito-temporal, le cortex cingulaire
antérieur, le cortex orbitofrontal,
avec comme autre caractéristique la
prédominance
hémisphérique droite»
(J Decety)
(4,7,8,27). Le cervelet participe
également par ses connexions avec les
noyaux gris centraux, comme agent de
coordination et de modulation de mouvements
automatiques innés ou acquis.
-
- Les études d'imagerie
cérébrale (4,8), chez l'homme,
s'accordent sur l'idée que la
perception des mouvements, des actions
réalisés par autrui, et
l'imagerie mentale de l'action partagent avec
la génération de l'action
intentionnelle un ensemble de régions
cérébrales (8,41). Elles
apportent des arguments en faveur de
l'hypothèse d'un codage commun
perception - action, compatibles avec la
notion d'affordance de Gibson (14) : «
Gibson désigne ainsi le fait que, dans
une situation donnée, l'environnement
se prête à une action
déterminée: une chaise incite
à la positon assise, un ballon attend
le coup de pied. Même si l'action n'est
pas exécutée, c'est en termes
d'action potentielle que l'environnement est
perçu. La perception des affordances
ne se limite pas aux seules relations avec
les objets inanimés et s'étend
à la dimension sociale contenue dans
l'environnement ». Le bâillement
est-il perçu comme une affordance
?
-
- Les travaux de J
Decety et de son équipe (8,27) ont
identifié les structures corticales
engagées lorsqu'un sujet imite les
actions réalisées par un
expérimentateur ou lorsque ses propres
actions sont imitées par
l'expérimentateur, comparé
à une condition de production
d'actions sans imitation : «Comme
attendu, en plus des régions
impliquées dans le contrôle
moteur, un réseau d'activations commun
au sein du cortex pariétal et du lobe
frontal (les régions
préfrontales dorso-médianes) a
été détecté entre
ces deux conditions d'imitation. Ce
réseau d'activations partagées
est cohérent avec l'hypothèse
d'un codage commun entre les actions du soi
et celles d'autrui. Si l'on regroupe les
données neurophysiologiques concernant
l'implémentation neuronale des trois
types d'activité (préparation,
simulation et observation pour imiter) qui
impliquent les représentations
motrices, on s'aperçoit qu'il existe
une étroite équivalence
fonctionnelle entre elles.»
-
- L'activation du gyrus frontal
inférieur dans
l'hémisphère gauche (qui
correspondrait à la région F5
chez le singe), au cours de l'observation
d'actions, peut s'expliquer par une
verbalisation silencieuse des sujets. Cette
zone appartient en effet à la
région de Broca (26) dont la
lésion provoque une aphasie de
production. Rizzolati
(39,40,41) a découvert dans le cortex
prémoteur ventral du macaque, dans les
aires F4 et F5, des groupes de neurones
appelés neurones miroirs dont
l'activité est corrèlée
à l'observation d'une action d'autrui
en fonction de son but, catégorisant
les actions à un niveau intentionnel.
Ces neurones semblent exister chez l'homme.
"The mirror system
would allow the observer to get into the
mental shoes of the target" V.
Gallese.
-
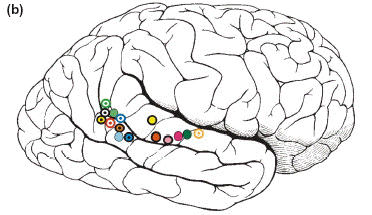
- En juin 2003, R.
Hari a présenté le premier
travail d'imagerie fonctionelle
cérébrale,
réalisé lors de la perception
du bâillemnt d'autrui. Le sillon
temporal supérieur, appartenant
à la zone des neurones miroirs,
s'active électicement lors de cette
perception et n'entre pas en fonction lors de
l'observation d'autres mouvements expressifs
d'un visage. Il s'agit là de la
première preuve scientifique d'un
mécanisme neurophysiologique
spécifique de la réplication du
bâillement.
- (Contagious
yawning: the role of self-awareness and
mental state attribution Platek SM)
-
- Percevoir les actions
réalisées par autrui
impliquerait un processus de simulation qui
permettrait d'en comprendre les intentions.
Cette résonance chez l'observateur ne
produit pas nécessairement un
mouvement ou une action. Un mécanisme
inhibiteur, parallèlement
activé et qu'on peut situer au niveau
frontal, bloquerait le déclenchement
moteur mimétique des actions. En
effet, l'étude de la pathologie
neurologique humaine des dysfonctionnements
frontaux (26) retrouve deux circonstances
où l'imitation non inhibée
perturbe les comportements:
- -1°) La
maladie de Gilles de la Tourette,
touchant le cortex préfrontal, les
ganglions de la base et le système
limbique, associe trois
éléments principaux: les tics,
la rare coprolalie, et
l'écholalie/échopraxie;
- -2°) Le
syndrome préfrontal ou
prémoteur associe une aphasie
kinétique (lésions
hémisphère gauche), et des
troubles de la sélectivité des
schémas moteurs alors que les
fonctions supérieures sont
respectées. (désautomatisation
des activités avec
persévérations et imitation
rudimentaire et erronée des derniers
mouvements de la personne face au malade,
cétopraxie).
- La réplication du bâillement
serait-il un comportement non inhibé,
de façon physiologique, et se
rapprochant par ses mécanismes de ces
pathologies ?
-
- A quel âge, chez l'homme, la
réplication du bâillement
apparaît-elle ? Piaget, dès
1951(29,32), avait montré que
l'imitation du bâillement par le
bébé n'apparait qu'au cours de
la deuxième année de vie, alors
que les nouveaux-nés bâillent
fréquemment, prolongeant ainsi ce
comportement apparu précocement au
cours de la vie foetale. Meltzoff
propose une interprétation
ontogénique à cette
discordance. Dans les six premiers mois de la
vie, le bébé est capable
d'imiter les mouvements des mains parce qu'il
voit les mains d'autrui comme les siennes.
Par contre, il n'a pas la perception
consciente de lui-même comme individu
ni perception des mouvements de son visage.
Lorsque le test du miroir indique qu'il a sa
propre perception d'individu autonome, qu'il
acquiert la capacité de se
reconnaître dans le miroir, son
développement mental de l'imitation
s'achève par la capacité
à imiter des mimiques, expliquant
ainsi qu'il devienne sensible à la
réplication du bâillement au
cours de la deuxième année de
vie seulement. L'unique étude
systématique des enfants indique,
elle, que cette réplication n'ait
observé qu'à près
l'âge de 5 ans (Anderson
& Meno 2003).
- F.
Giganti et al. ont montré que la
réplication du bâillement
était une capacité qui
s'altérait avec l'âge,
très réduite chez les personnes
très âgées. Cette
évolution est similaire et
parallèle à la réduction
des performances attachées à la
théorie de l'esprit.
-
- La
réplication du bâillement chez
les chimpazés
-
- Bien que le bâillement soit
répandu chez tous les
vertébrés, sa
réplication n'a été
rapportée jusqu'alors que chez les
humains (Lehmann
1979; Baenninger
1987; Smith 1999).
Cette capacité est sous-tendue par les
mécanismes neuropsychologiques de la
théorie de l'esprit (Theory of Mind) ,
puissant moyen de communication
inter-indiviudelle à la base de
l'empathie. Un travail récent a ainsi
pû montrer que cette capacité
à subir la réplication
était d'autant plus nette que la
personnalité est empathique par
opposition aux personnalités
schizoïdes très peu sensibles
(Platek et
al. 2003). L'équipe de recherche
dirigée par James Anderson de
l'Université de Stirling (GB) a , pour
la première fois, réussi
à retrouver cette capacité
à la réplication des
bâillements chez les chimpanzés,
nos proches parents
phylogénétiques, lorsqu'ils
regardent un enregistrement vidéo d'un
congénère qui bâille.
Cette découverte est un pas de plus
pour montrer que nous partageons avec des
primates non-humains des traits
évolutifs communs pour la
capacité à percevoir la
conscience de soi-même et de celle des
autres. Le
mâle dominant fait plus bâiller
les autres, c'est à dire que les
autres sont plus sensibles aux
bâillement de leur 'patron'. Chez
les bonobos, singes sociaux dont le
rôle dominant appartient aux femelles,
les apparentés sont plus sensibles,
puis les proches.
-
- La photo ci-dessous montre le reflet de
l'écran vidéo où se
déroule le bâillement d'un
chimpanzé et visualise le
bâillement du chimpanzé
spectateur. Voir
cette vidéo.
-
 -
- Il semble que, sous certaines conditions
de proximité et complicité
anciennes, entre maître et chien,
le
chien puisse être sensible aux
bâillements de son
maître.
-
- En résumé, admettant
que le développement du cortex frontal
(moteur) et préfrontal
(prémoteur) est spécifique aux
bipèdes, on peut donc proposer que la
réplication du bâillement est
une véritable
échokinésie, pour reprendre ce
mot inventé par JM Charcot, que l'on
peut caractériser par trois
critères :
- -la réplication serait une
spécificité humaine
interprétée comme un
mimétisme comportementale
- -alors que l'observation d'un
comportement moteur d'autrui est mimée
par les aires motrices de l'observateur et le
plus souvent non suivi d'actes moteurs par
inhibition frontale, le bâillement
serait-il, lui, sous certaine condition de
niveau de vigilance, le résultat d'un
comportement non inhibé ?
- -la réplication aurait
conféré un avantage
sélectif en permettant une
synchronisation efficace des niveaux de
vigilance entre les membres d'un groupe. Elle
participerait d'une forme d'empathie
instinctive involontaire, probablement
apparue tardivement au cours de
l'évolution des hominidés.
-
- Archaic
behavior and the communicative act
Meerloo J 1955 et
pdf
-
- Imitation et intersubjectivité :
voir un site de l'INSERM Prof Jean Decety
(PETscann et IRM fonctionnelle)
- lire les articles
de J Decety sur ce site
-
- Théorie
de l'esprit, empathie et...
bâillement C Derouesné
-
- Contagious
yawning: the role of self-awareness and
mental state attribution Platek SM, SR
Critton, et al
- Yearning to
yawn: the neural basis of contagious
yawning Schurmann M, Hari R et al
- Contagious
yawning and the brain Platek S, Mohamed
F, Gallup G
- Cortical
systems for the recognition of emotion in
facial expressions R Adolphs, A
Damasio
- Human
facial expressions as adaptations:
evolutionary questions in facial expression
research KL Schmidt
- Neurophysiological
mechanisms underlying the understanding and
imitation of action G Rizzolati, V
Gallese
- From
the perception of action to the understanding
of intention J Decety
- Imitation and
the definition of a meme Susan
Blackmore
- La
communication chez les primates BL
Deputte
- Imiter
pour découvrir l'humain J Decety
et J Nadel
- A PET
exploration of the neural mechanisms involved
in reciprocal imitation J Decety, T
Chaminade, J Grèzes, AN Meltzoff
- Is
perceptual anticipation a motor simulation ?
A PET study T Chaminade, D Meary, JP
Orliaguet, J Decety
- Does
the End Justify the Means? A PET Exploration
of the Mechanisms Involved in Human
Imitation T Chaminade, AN. Meltzoff, J
Decety
- Naturaliser
l'empathie J Decety
- Neural
correlates of feeling sympathy J Decety
et T Chaminade T
- Functional
imaging of theory of mind: the role of the
STS Gallagher HL
- Resonance
behaviors and mirror neurons G
Rizzolatti, V Gallese
- Reafferent
copies of imitated actions in the right
superior temporal cortex M Iacoboni
- Imitation of
facial and manual gestures by human
neonates Meltzoff AN, Moore MK
- Rational
imitation in preverbal infants G
Gergely
- Imitation
et agentivité J Proust
- La
communication chez les primates BL
Deputte
- The
perception-behavior expressway:automatic
effects of social perception on social
behavior
- The
temporal relationship between reduction of
early imitative responses and the development
of attention mechanisms Nakagaw A
- A
clinical and psychological study of
echo-reactions Srengel Ed, Vienna MD,
Edin LR 1947
-
- Un
précurseur des recherches actuelles...
oublié ! Yawning,
Yielding, and Yearning to
Yawn
- Paper presented at the
meeting of the Midwest Psychological
Assiociation Chicago,
May 1974
- Cialdini
RB, McPeek RW
-
- -Joly-Mascheroni
RM, Senju A, Sheperd AJ Dogs catch human
yawns Biology letters Animal Behaviour
2008;4(5):446-448
- -Nahab
FB, Hattori N, Saad ZS, Hallett M
Contagious yawning and the frontal lobe: An
fMRI study. Human Brain Mapping
2009;30:1744-1751
-
- Empathy
in schizophrenia: impaired resonance
Haker H, Rössler W
 -
 -
    -
-
-
-
- -Arnott
SR et al. An investigation of auditory
contagious yawning Cognitive, Affective,
Behavioral Neurosci 2009;9(3):335-342
- -Anderson
JR et al Contagious yawning in
chimpanzees The Royal Society Biology Proc R
Soc Lond B Biol Sci 2004; 271 Suppl 6;
S468-470
- -Anderson
JR et al Psychological influences on
yawning in children Current Psychology
Letters Behaviour, Brain, Cognition
2003;2:11
- -Anderson
JR, Matsuzawa T. Yawning: An Opening into
Empathy? Cognitive Development in
Chimpanzees. T Matsuzawa, M Tomonaga, M
Tanaka Editors. Springer 2006
- -Caswell
TA, The effects of status on yawning
behavior. Thesis 1991
- -Cialdini
RB, McPeek RW. Yawning, Yielding, and
Yearning to Yawn 1974 (non published)
- -Campbell
M et al. Computer animations stimulate
contagious yawning in
chimpanzees Proceed Royal Soc Biol
2009:276(1676):4255-4259
- -Campbell
MW, de Waal FB. Ingroup-Outgroup Bias in
Contagious Yawning by Chimpanzees Supports
Link to Empathy. Plos One. 2011;6(4):1-4
- -Cooper
NR, Puzzo I, Pawley A Contagious yawning:
the mirror neuron system may be a candidate
physiological mechanism Medical Hyportheses
2008;71(6):975-976
- -Cooper
NR, Puzzo I, et al. Bridging a yawning
chasm: EEG investigations into the debate
concerning the role of the human mirror
neuron system in contagious yawning.Cogn
Affect Behav Neurosci.
2012;12(2):393-405
- -Demuru
E, Palagi E. In Bonobos Yawn Contagion Is
Higher among Kin and Friends. PLoS One. 2012;
7(11): e49613
- -Derouesné
C. Théorie de 'esprit, empathie
et... bâillement. Psychologie &
NeuroPsychiatie du vieillissement
2003;1(4):1
- -de
Wall F. L'âge de l'empathie. Les
liens qui libèrent. 2010:75-86
- -Dijksterhuis
A, Bargh JA The perception-behavior
expressway:automatic effects of social
perception on social behavior Advances in
Experimental Social Psychology
2001;33:1-40.
- -Estow,
S Jamieson JP, Yates JR Self-monitoring
and mimicry of positive and negative social
behaviors Journal of Research in Personality
2007;41(2):425-433
- -Gallagher
HL, Frith CD Functional imaging of theory
of mind Trends in Cognitive Scie.
2003;7(2):77-83
- -Giganti
F, Ziello ME Contagious and spontaneous
yawning in autistic and typically developing
children CPL 2009
- -Giganti
F, Zilli I. The daily time course of
contagious and spontaneous yawning among
humans. J Ethol 2011;29(2):215-216
- -Giganti
F, Toselli M, Ramat S. Developmental
trends in a social behaviour: contagious
yawning in the elderly. Giornale di
Psicologia dello Sviluppo.
2012;101:111-117
- -Hagenmuller
F, Rössler W, Endrass J, Rossegger A,
Haker H.Impaired resonance in offenders
with psychopathic traits Neuropsychiatr.
2012;26(2):65-71. article in german
- -Haker
H, Kawohl W, Herwig U, Rössler W.
Mirror neuron activity during contagious
yawning-an fMRI study. Brain Imaging Behav.
2012
- -Haker
H, Rössler W.
Empathy in schizophrenia: impaired
resonance. Eur Arch Psychiatry Clin Neurosci
2009;259(6):352-361
- -Harr
AL, Gilbert VR Do dogs show contagious
yawning ? Anim Cogn. 2009;12(6):833-837
- -Helt
MS, Eigsti IM, Snyder PJ, Fein DA.
Contagious yawning in autistic and typical
development. Child Dev.
2010;81(5):1620-1631
- -Iacoboni
M, G Rizzolatti Cortical mechanisms of
human imitation Science 24/12/99; 286
- -Joly-Mascheroni
RM, Senju A, Sheperd AJ Dogs catch human
yawns Biology letters Animal Behaviour
2008;4(5):446-448
- -Lindsay
SR Coping with fear and stress: licking
and yawning. Handbook of applied dog behavior
and training 2000
- -Madsen
EA, Persson T. Contagious yawning in
domestic dog puppies (Canis lupus
familiaris): the effect of ontogeny and
emotional closeness on low-level imitation in
dogs. Anim Cogn. 2012
- -Massen
JJ, Vermunt DA, Sterck EH. Male Yawning
Is More Contagious than Female Yawning among
Chimpanzees (Pan troglodytes). PLoS One.
2012;7(7):e40697.
- -Millen
A, Anderson JR Neither infants nor
toddlers catch yawns from their mothers. Biol
Letters 2011;7(3):440-442
- -Miller
ML, Gallup AC, Vogel AR, Vicario SM, Clark
AB. Evidence for contagious behaviors in
budgerigars (Melopsittacus undulatus): an
observational study of yawning and
stretching. Behav Processes. 2012
Mar;89(3):264-270
- -Nahab
FB, Hattori N, Saad ZS, Hallett M
Contagious yawning and the frontal lobe: An
fMRI study Human Brain Mapping
2009;30:1744-1751
- -Norscia
I, Palagi E. Yawn Contagion and Empathy
in Homo sapiens. PLoS ONE. 2011;6(12):
e28472
- -O'Hara
SJ, Reeve AV A test of the yawning
contagion and emotional connectedness
hypothesis in dogs, Canis familiaris. Animal
Behaviour 2011;81:335-340
- -Palagi
E, Leone A, Mancini G, Ferrari PF.
Contagious yawning in gelada baboons as a
possible expression of empathy. Proc Natl
Acad Sci USA. 2009;106(46):19262-19567
- -Paukner
A, Anderson JR
Video-induced yawning in stumptail
macaques (Macaca
arctoides) Biol Lett 2006;2(1):36-38
- -Perkins
JR Teaching Dogs to Yawn, Sneeze, and
Implications for Preparedness Theory and
Observational Learning.
In: Kusonose, Ryo and Sato, Shusuke
39th Congress of the International Society
for Applied Ethology, Kanagawa, Japan. 20-24
August, 2005
- -Platek
SM, SR Critton, et al Contagious yawning:
the role of self-awareness and mental state
attribution Cogn Brain Res
2003;17(2):223-227
- -Platek
S et al. Neural correlates of self-face
recognition 2008 Brain Res;1232:173-184
- -Platek
S, Mohamed F, Gallup G Contagious yawning
and the brain Cognitive Brain Research,
2005;23:448-452
- -Schurmann
M, Hari R et al Yearning to yawn: the
neural basis of contagious yawning NeuroImage
2005;24(4):1260-1264
- -Senju
A, Maeda M, Kikuchi Y, Hasegawa T, Tojo Y,
Osanai H Absence of contagious yawning in
children with autism spectrum disorder.
Biology Letters 2007;3:706-708
- -Senju
A, Kikuchi Y, Akechi H et al. Does eye
contact induce contagious yawning in children
with autism spectrum disorder? J of Autism
and Developmental Disorders.
2009;39(11):1598-1602
- -Silva
K, Bessa J, de Sousa L. Auditory
contagious yawning in domestic dogs (Canis
familiaris): first evidence for social
modulation. Anim Cogn. 2012.
- -Yoon
JMD, Tennie C Contagious yawning: a
reflection of empathy, mimicry, or contagion.
Anim Behav 2010;79:e1-e3
- -Walusinski
O Echokinetic yawning, theory of mind,
and empathy
- -Wilkinson
A et al. No evidence of contagious
yawning in the red-footed tortoise. Current
Zoology 2011;57(4):477-484
-
- by Pedro Forza

|