- The central administration of
adrenocorticotropin (ACTH) and
a-melanocyte stimulating hormone
(alpha-MSH) induces in different animal
species a complex behaviour characterized by
recurrent episodes of yawning, stretching,
penile erection and ejaculation.
- This behaviour is considered a specific
central effect of ACTH-MSH peptides, since it is
not observed after their peripheral
administration, nor is induced by any other
known peptide tested (1-5). It was found that
the total peptidic extract from few rat
hypothalami produced penile erection.
ejaculation, yawning and stretching when
injected into the lateral ventricle of a
recipient rabbit (3). This finding is surprising
because the total hypothalamic content of the
combination ACTH and a-MSH is about a thousand
times lower than the minimum dose of these
peptides necessary to induce the above mentioned
effects (6,7). This raises the possibility that
some other substance, able to induce these
behaviours, might be present in the hypothalamus
in addition to ACTH and a-MSH. In order to test
this hypothesis, we fractioned a peptidic
extract from rat hypothalamus by high pressure
liquid chromatography (HPLC), and tested each
fraction for its ability to induce yawning.
stretching and penile erection. Here we report
that another peptide is present in the
hypothalamus which is capable of induping penile
erection and yawning with a potency of at least
500 times that of ACTH and a-MSH. This peptide
is oxytocin.
Among the 45 fractions tested, only that
eluting at 15 min induced penile erection and
yawning when injected into the recipient
animals. The equivalent of 15 hypothalamic
induced at least 4 penile erections and 20 yawns
during the 60 min observation. Biological
activity was destroyed by digestion with Pronase
(Boehringer Kannheim) indicating that the active
substance was a peptide(s). In order to identify
the active peptide, the active fraction from
1000 hypothalami was combined after the first
HPLC step, reinjected in the Bondapak C18
colomn, and eluted isocratically with 22% B. The
activity eluted es a sharp peak at 13 min,
either at 210 or 280 nm. Amino acid and sequence
analysis, performed as previously described
(10), revealed that the peptide was identical to
oxytocin.We calculated from amino acid anilysis
that 6 nmol of oxytocin were isolated from 1000
hypothalami. Indeed synthetic oxytocin coeluted
with the hypothalamic peptide.
The ability of oxytocin to induce penile
erection and yawning was unexpected since in
previous studies oxytocin was found completely
inactive(3). These contrasting observations led
us to further characterize oxytocin effect on
penile erection and yawning. A significant
increase in the number of penile erections and
yawns occurred when a dose as low as 5 ng of
oxytocin was injected into a lateral ventricle.
Yawning episodes, alone or together with
stretching, usually started 8-10 min after
i.c.v. injection, and the highest number of
yawns was seen with the dose of 20 and 60 ng of
the peptide. These doses also induced the
highest number of penile erections and intense
genital grooming. The symptomatology lasted for
1 to 2h. However, in contrast to low doses, the
injection of oxytocin in doses of 120 and 1200
ng produced neither yawning nor penile
erection.
Unlike oxytocin, the i.c.v.injection of
equimolar doses of arg-vasopressin, which
differs from oxytocin only in 2 amino acids,
Phe3 and Arg8 instead of ile3 and Leu8 failed to
induce sexual stimulation and yawning. In
agreement with previous studies (11), doses of
arg-vasopresssin higher than 5 ng caused severe
motor disturbances and barrel rotation, a
symptonatology that lasted for 15-20 min after
treatment. After recovery the animals showed an
apparently normal behaviour but failed to show
yawning or penile erection for up to 1 hr.
Equimolar amounts of i.c.v. ACTH 1-24. a-MSH,
corticotropin releasing factor (CRF), delta
sleep inducing peptide, neurotensin and
substance P produced no-overt behavioural
changes. ACTH 1-24 and a-MSH induced stretching,
yawning and penile erection at doses of 2 nmol
(results not shouwn), in agreement with previous
studies (1-3).
In order to clarify the mechanism of
oxytocin-induced penile erection and yawning, we
studied the influence of different drugs on this
behaviour. Pretreatment with antimuscarinic
atropine (10 mg/kg i.p.) or morphine (5 mg/kg
i.p.) completely abolished penile erection and
yawning induced by the peptide. On the contrary,
methylatropine (10 mg/kg i.p.), that does not
cross the blood brain barrier, and the opiate
antagonist naloxone (l mg/kg i.p.) were
ineffective. Haloperidol (l mg/kg i.p.), a
dopamine receptor antagonist, prevented sexual
behaviour, but not yawning.
The present results show that the
intraventricular injection of very low doses of
oxytocin induces penile erection and yawning in
male rats. No explanation is available at
present for the ineffectiveness of relatively
high doses of the peptide to induce such
behaviours, although this is in agreement with
previous studies showing an inverted U-shaped
dose-response curve for other effects of
oxytocin (12,13).
The ability by different drugs to interfere
with oxytocin-induced penile erection and
yawning, gives an indication of the complex
central mechanisms underlying such behaviours.
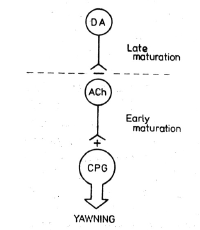
Atropine prevention of oxytocin effects
indicates that the central cholinergic system
plays a role in the expression of penile
erection and yawning in agreement with previous
studies (14). Similarly, prevention by morphine
of penile erection and yawning suggests that
central opioid systems are implicated in the
expression of such behaviours induced by
oxytocin. However it is unlikely that opioids
exert a tonic inhibitory control. on such
behaviours since opioid receptor blockade by
naloxone was ineffective in modifying oxytocin
effect. On the other hand, central dopaminergic
systems seen to be implicated mainly in the
expression of the sexual response to oxytocin
since the latter effect but not yawning was
antagonized by dopamine receptor blockade
suggesting that the two behaviours might be
mediated by different mechanisms, in spite of
the fact that they often appear concomitantly.
Accordingly, castration has been shown to
eliminate the sexual response, but leaving the
yawning response to ACTH-MSH peptides unmodified
(2,3).
So far only two means were known to be able
to induce both penile erection and yawning in
experimental animals: one is the systemic
injection of apomorphine and other
dopaminomimetic agents (15-17), the second is
the injection of ACTH-MSH peptides into the
cerebrospinal fluid or specific brain areas
(2,3). We have shown that oxytocin also induces
these behaviours. Whether or not penile erection
and yawning induced by oxytocin, ACTH-MSH
peptides and dopaminomimetic drugs, are mediated
by the same mechanisms (i.e. endogenous oxytocin
is implicated in the expression of such
behavicurs in:juced by ACTH-MSH peptides and
dopaminergic drugs or vice-versa), is unknown at
present.
-
- Indeed oxytocin effects are antagonized by
atropine and morphine, but not by naloxone,
similarly to ACTH-MSH- and
dopaminomimetic-induced yawning and sexual
behaviour (1,2,14). However, two important
features distinguish oxytocin effect from that
of ACTH-MSH peptides: 1) the doses of oxytocin
that induce penile erection and yawning (5.-60
pmol) are much lower than those of ACTH-MSH
peptides (1-2 pmol) (2,3); and 2)
oxytocin-induced penile erection and yawning
be&in 8-10 min afte.-.i.c.v. injection of
the peptide, while a lag of 25-30 min is always
observed after i.c.v. injection of ACTH-MSH
peptides (2,3). The reason for the long latency
in the effect of ACTH-MSH peptides has never
been clarified. It is tempting to speculate that
ACTH-MSH peptides induce yawning, stretching and
sexual behaviour by releasing oxytocin in some
brain area. The latency in ACTH-MSH response
might be due to the time necessary for ACTH-MSH
peptides to release enough oxytocin to elicit
the symptomatology.
-
- Similarly dopaminomimetic drugs might induce
yawning and penile erection by enhancing
oxytocin release in some brain area. According
to this hypothesis, experimental evidence
indicates that oxytocin may act as a
neuropeptide. Indeed oxytocin is synthetized in
neurons originating in the hypothalamus that
send their projections not only to the
neurohypophysis but elso to other brain areas,
such as amygdala, frontal cortex, septum,
hippocampus, pons and medulla (18-20), with an
innervation independent from that of the other
neurohypophyseal peptide vasopressin (21-23),
and an oxytocinergic receptor has been
characterized in rat brain (24). Furthermore
oxytocin has been implicated in the modulation
of memory processes (11,25-27), in the induction
of maternal behaviour (28), in the development
of narcotic tolerance and physical dependence
(13), and may be the precursor of patent
biologically active neuropeptides (29).
In conclusion, oxytocin is the most patent
agent discovered sa far that produces penile
erection and yawning. Although the mechanism and
the site(s) of action of the peptide responsible
for the induction of such behaviours are
unknown, the present results suggest a
physiological role of oxytocin in the expression
of Bexual behaviour and yawning, and provide
further evidence that oxytocin, in addition ta
its hormonal rolc in parturition and lactation,
acts as a neuropeptide in the central nervous
system.
References
- W. Ferrari, G.L. Gessa and L. Vargiu, Ann.
N.Y. Acad. Sci. 104, 330 (1963).
- A. Bertolini and G.L. Gessa, J. Endocrinol.
Invest. 4, 241 (1981).
- A. Bertolini, G.L. Gessa and W. Ferrari, in
Sexual Behaviour - Pharmacology and Biochenistry
(M. Sandler and G.L. Gessa eds.) pp. 247, Raven
Press, New York (1975).
- T.L. O'Donohue and D.M. Dorsa, Peptides 3,
353 (1982)
- A.J. Kastin, N.P. Plotnikoff, A.V. Schally
and D.H. Coy, in Reviews of Neuroscience (S.
Ehrenpreis and I.J. Kopin, eds.) Raven Press,
New York (1976).
- D.T. Krieger, A. Liotta and MJ. Brownstein,
Proc. Natl. Acad. Sci. U.S.A. 74, 648
(1977).
- A. Argiolas, MR. Melis, W. Fratta and G.L.
Gessa, Peptides
- L. Pellegrino and A.J. Cushman, Meredith,
New York (1971).
- H.P.J. Bennett, C.A. Browne and S. Solomon,
Biochemistry 20, 4530 (1981).
- A. Argiolas A. and J.J. Pisano, J. Biol.
Chem. 260, 1437 (1985).
- H. Kruse, TB. Van Wimersma Greidenus and D
de Wied, Pharmacol. Biochem Behav. 7, 311
(1977).
- B. Bohus, I. Urban, Tj.B. Van Wimersma
Greidanus and D. de Wied, Neuropharmacology 17,
239 (197b).
- G.L. Kovacs, Z. Horvath, Z. Saruyai, M.
Faludi and G. Teledgy, Neuropharnacology 24, 413
(1985).
- K. Yamada and T. Furukara,
Psychopharmacology 67, 39 (1980).
- E. Mogilnicka
and V. Klimek, Pharmacol. Biochem. Behav. 7.
303 (1977).
- G. Serra, W. Fratta, M. Collu, L.
Napoli-Farris and G.L. Gessa, Eur. J. Pharmacol.
86. 279 (1983).
- C.T. Dourish, S.J. Cooper and C.R. Philips,
Psychopharmacology 86, 175 (1985).
- R.M Buijs, Cell Tissue Res. 192, 423
(1978).
- M.W. Sofroniew and U. Schrell, Neuroscience
Lett. 22, 211 (1981).
- G. Valiquette, J. Haldar, G.M Abrams, G.
Nilaver and E.A. Zimmermann, Brain Res. 331, 176
(1985).
- G.J. de Vries and R.M. Buijs, Brain Res.
273, 307 (1983).
- R.E. Lang, J. Heil, D. Ganten, K. Herman. W.
Rascher and Th. Unger, Brain Res. 260, 326
(1983).
- MW. Sofroniew, Neuroscience 15, 347
(1985).
- M Muhlethaler, W.H. Sawyer, M.M. Manning and
J.J. Deifruss, Proc. Natl. Acad. Sci.U.S.A. 80,
6713 (1963).
- J.M. VLri Ree, B. Bohus, D.H.G. Versteeg and
D. de Wied, Biochem. PharmecDI. 27, 1793
(1976).
- D. de Wied, Proc. Roy. Soc. London B210, 183
(1980).
- R. Walter, P.L. Hoffman, J.B. Flexner and
L.B. Flexner, Proc. Natl. Acad. Sci. U.S.A. 72,
4180 (1975).
- C.A. Pedersen, J.A. Ascher, Y.L. Monroe and
A.J. Prenge, Science 216, 648 (19E2).
- JP.H. Burbach., B. Bohus, G.L. Kovacs, J.M.
Van Nispen, H.M. Greven and D. de Wied, Eur. J.
Pharmacol. 94, 125 (1983).
|