It has been generally accepted that
cerebrospinal fluid (CSF), bathing the central
nervous system, is mainly produced within the
cerebral ventricular system, and circulates as a
slow river from the brain ventricles towards the
cortex subarachnoid space to be absorbed across
the arachnoid villi into the venous
sinuses.
From time to time, this hypothesis bas been
challenged on the basis of some experimental
data. However, in spite of the incompatibility
of differences between the mentioned hypothesis
and these experimental results, the hypothesis
still persists in the unchanged form. Namely,
the methods applied to determine the way of CSF
formation and circulation have been those that
provide indirect inspection of the
aforementioned processes by following changes in
the characteristics or behavior of a substance
after its application into the CSF system. For
example, dilution of substances applied in CSF
has most frequently been used as a measure of
the CSF formation while the distribution of
(labelled) substances all over the CSF system
has most frequently been used as a marker of the
CSF circulation. So, any mistake in the
interpretation of the dilution (escape of
substances into the brain tissue, the mixing
problem, etc.) or distribution (molecular
weight; the site and way of application, etc.)
of substances in CSF will result in questionable
and often contradictory conclusions regarding
the CSF formation and circulation.
To avoid the indirect study of the CSF
formation and circulation we developed a new
experimental model in which these two
physiological parameters were examined under
direct vision. The first attempt of a direct
vision study was shown in our preliminary study.
That attempt consisted of a surgical approach
with poor control of the CSF and atmospheric
pressure. For this reason, in new model,
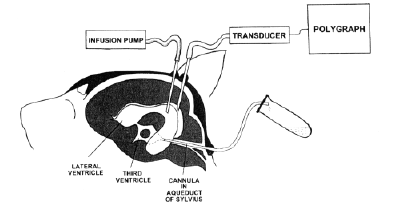
presented in this paper, the aqueduct of Sylvius
of anaesthetized cat was cannulated by a plastic
cannula and surgical reconstruction was done so
that the CSF system was completely protected
against any uncontrolled influences of
atmospheric pressure and CSF leakage from
subarachnoid and ventricular spaces. Thus. the
relationship between the ventricular and
subarachnoid CSF pressures was established in a
physiological range without a transmantle
pressure gradient which is a crucial advantage
of our model. Namely, in case of pathway
obstruction, the absence of a transmantle
gradient would not cause a possible CSF flow
from the ventricles through the brain tissue
into subarachnoid space, which was suggested by
Milchorat. In addition to that, the CSF pressure
control is also important because it has been
shown that the CSF formation is a pressure
dependent process.
If CSF is mainly formed inside the brain
ventricles and absorbed in the subarachnoid
space, it has to circulate at a physiological
CSF pressure through the aqueduct of Sylvius or,
as in our model, through the plastic cannula
positioned in the aqueduct. Therefore, the
direct (visual) observation of the CSF outflow
throughout the external end of the cannula,
adjusted to the physiological CSF pressure,
should represent the direction of the CSF
circulation from the ventricles to the
subarachnoid space. The collected volume of CSF
divided by the time of collection should
represent the rate of the CSF formation.
According to our previous results, where the CSF
formation in cat's brain ventricles was measured
indirectly by the dilution of blue dextran in
CSF and the rate of formation was calculated by
the equation of Heisey et al, the rate of
formation at least 4.5µl/ min should be
expected.
[...] These findings clearly
indicate that, in case of the experimentally
produced CSF formation, the CSF circulation was
established and CSF moved freely through the
ventricular system, from the site of infusion
(lateral ventricle) via the aqueduct of Sylvius
to the external end of the plastic cannula, with
the velocity of the circulation being equal to
the rate of infusion. Also, neither the
absorption nor the net formation of CSF were
observed inside the brain ventricles, because
neither the reduction or the enhancement outflow
were detected.The obtained results apparently
indicate that no CSF formation exists inside the
brain ventricles, and consequently that CSF does
not circulate through the brain ventricles at
all.
The presented data largely correspond to
the new hypothesis suggested by Bulat which
starts from the assumption that hydrostatic and
osmotic forces operate between the CSF volumes
in the way similar to the way they regulate the
volume of extra-cellular fluid in other
organs. Hydrostatic and osmotic forces seem
to play the main role in the regulation of the
CSF volume according to this new hypothesis, and
it seems that the imbalance between these forces
will result in changes in the CSF volume.
[...] In conclusion, in spite
of the fact that in our model animals were under
anaesthesia and that the fourth ventricle was
excluded from the investigation, the classical
hypothesis of the CSF formation and circulation
cannot be upheld, because our results clearly
show that the net CSF formation does not take
place inside the other brain ventricles and that
CSF does not circulate as a slow river from the
ventricles to the subarachnoid space.
It is very well known (and commonly applied
in routine clinical practice) that the increase
of blood osmolality can decrease the CSF
(intracranial) pressure, simply by extraction of
fluid from nervous tissue. Our experimental
results clearly revealed that the increase in
CSF osmolality will subsequently lead to the
increase of the CSF volume inside the brain
ventricles. This drives to a conclusion that
the osmolality represents one of the major
determinants of fluid exchange in intracranial
pressure. It seems that the control of the
CSF volume is under the influence of hydrostatic
and osmotic forces between the CSF system and
the surrounding tissue and that the CSF volume
will be changed, depending on the prevalence of
those forces, caused by (patho)physiological
reasons inside or outside the CSF system.
Anyhow, the results presented in this article
call for a new approach to the physiology and
pathology of CSF and we feel that time has come
to re-evaluate the classical hypothesis of the
CSF hydrodynamics to abandon this hypothesis as
the traditional framework of thinking.
-Legendre
R, Piéron H.
De la propriété
hypnotoxique des humeurs
développée au cours d'une veille
prolongée C.R. Société de
Biologie de Paris 1912; 70; 210-212
-Bouyssou M,
Tricoire J Experimental detection of a
cervical arousal mechanism of yawning, enhancing
intracerebral (CSF) fluid pressure J Dental Res
1985; 64; 721
-Lepp
FH Remarques sur la signification
physiologique du bâillement Bull Group Int
Rech Sci Stomtol Odontol 1982; 25; 251-290
-Nolman
B Yawning, cerebral fluid and the lymphatic
pump 2006
-Oreskovic D;
Klarica M; Vukic M The formation and
circulation of cerebrospinal fluid inside the
cat brain ventricles : a fact or an illusion ?
Neuroscience letters 2002; 327; 103-106
-Patra P, Gunness
TK, Robert R Physiologic variations of the
internal jugular vein surface, role of the
omohyoid muscle, a preliminary echographic study
Surg Radiol Anat 1988; 10; 2; 107-12
-Schniter E The
evolution of yawning : why do we yawn and why is
it contagious ? thèse 2001
-Schroth G, Klose U
Cerebrospinal fluid flow; Physiology of
respiration-related pulsations. Neuroradiology
1992; 35; 1; 10-15
-Walusinski
O Prostaglandines, adénosine, sommeil
& bâillements 2004
-Woodbury R, B
Abretj Influence of dying gasps, yawns and
sighs on blood pressure and blood flow Am J
Physiol 1944; 142; 721-726